

Cyclin-dependent kinase 5-deficient mice demonstrate novel developmental arrest in cerebral cortex
- PMID: 9698328
- PMCID: PMC6793186
- DOI: 10.1523/JNEUROSCI.18-16-06370.1998
Cyclin-dependent kinase 5-deficient mice demonstrate novel developmental arrest in cerebral cortex
Abstract
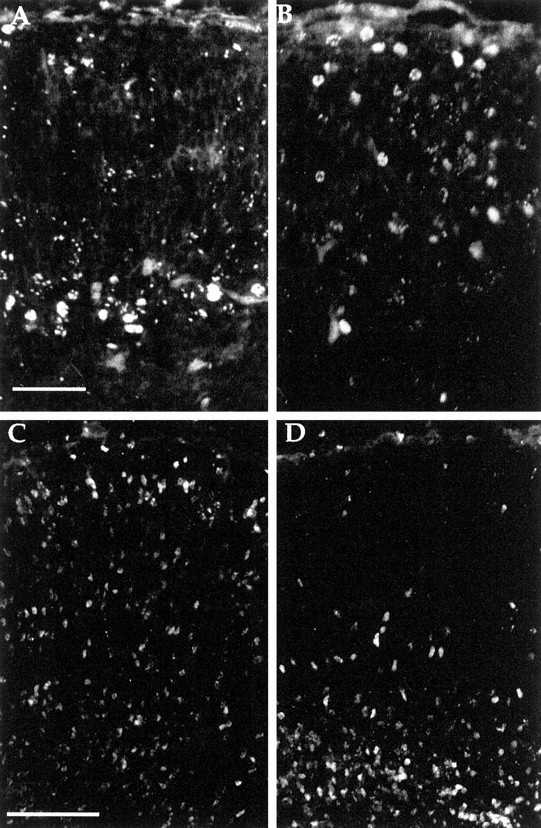
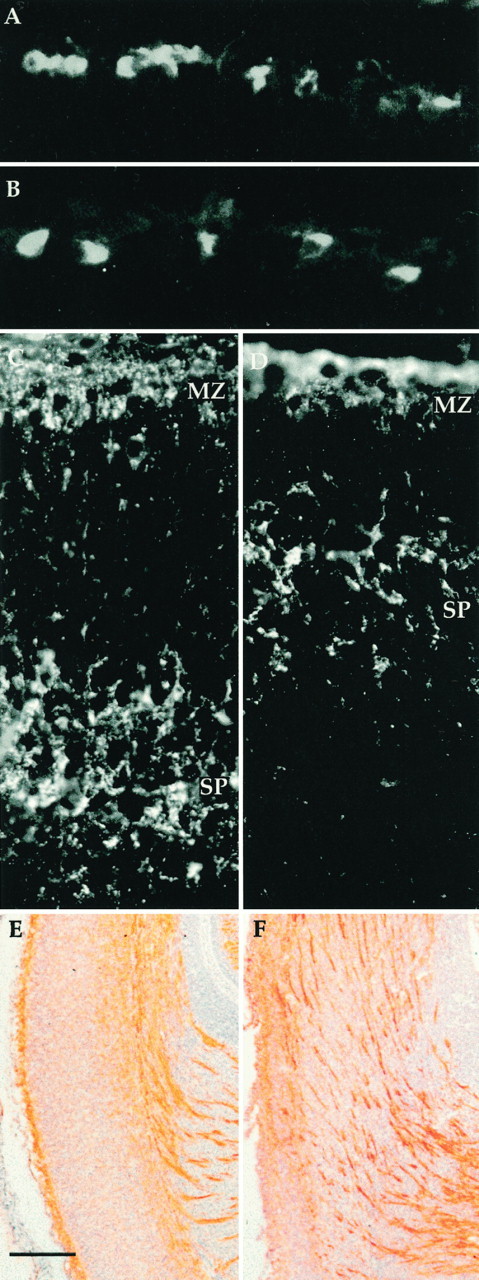
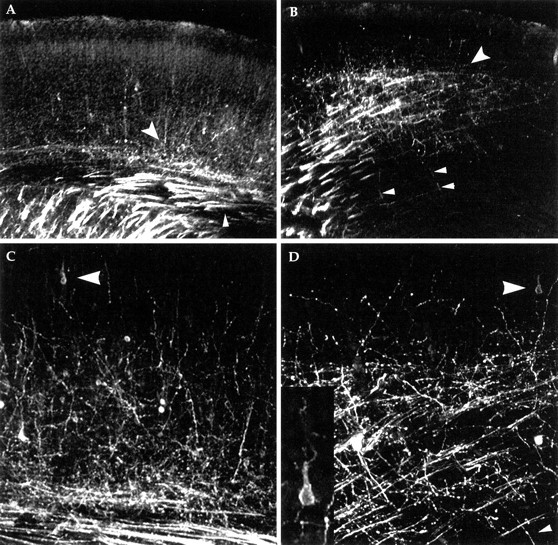
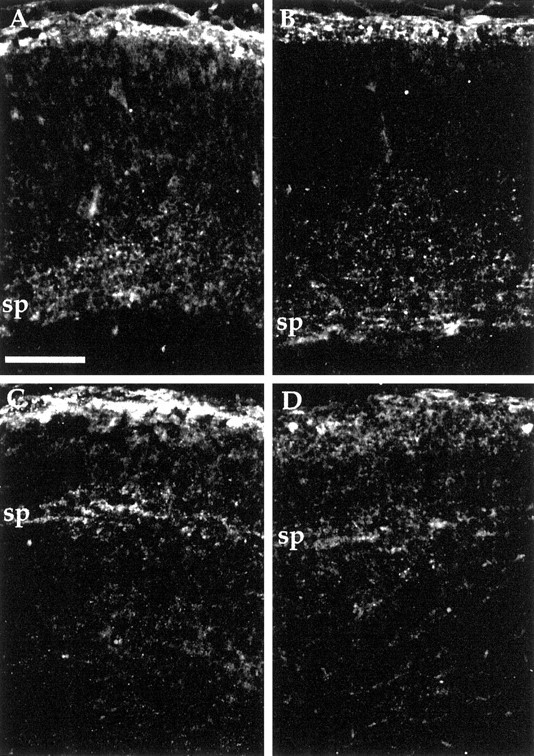
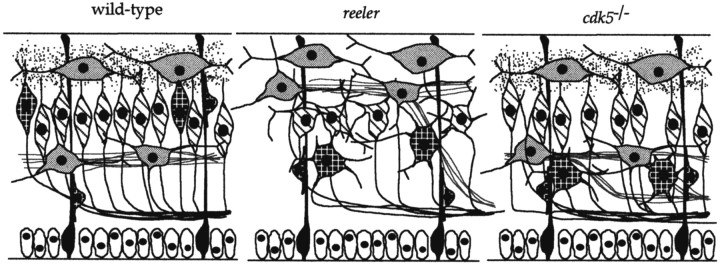
The cerebral cortex of mice with a targeted disruption in the gene for cyclin-dependent kinase 5 (cdk5) is abnormal in its structure. Bromodeoxyuridine labeling reveals that the normal inside-out neurogenic gradient is inverted in the mutants; earlier born neurons are most often found superficial to those born later. Despite this, the early preplate layer separates correctly and neurons with a normal, pyramidal morphology can be found between true marginal zone and subplate. Consistent with their identity as layer VI corticothalamic neurons, they can be labeled by DiI injections into thalamus. The DiI injections also reveal that the trajectories of the cdk5(-/-) thalamocortical axons are oblique and cut across the entire cortical plate, instead of being oriented tangentially in the subcortical white matter. We propose a model in which the cdk5(-/-) defect blocks cortical development at a heretofore undescribed intermediate stage, after the splitting of the preplate, but before the migration of the full complement of cortical neurons.
Figures

References
-
- Allendoerfer KL, Shatz CJ. The subplate, a transient neocortical structure: its role in the development of connections between thalamus and cortex. Annu Rev Neurosci. 1994;17:185–218. - PubMed
-
- Angevine JB, Sidman RL. Autoradiographic study of cell migration during histogenesis of cerebral cortex in mice. Nature. 1961;192:766–768. - PubMed
-
- Boulder-Committee. Embryonic vertebrate CNS: revised terminology. Anat Rec. 1970;166:257–261. - PubMed
-
- Brittis P, Meiri K, Dent E, Silver J. The earliest patterns of neuronal differentiation and migration in the mammalian central nervous system. Exp Neurol. 1995;134:1–12. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases