

Synchronization of visual responses between the cortex, lateral geniculate nucleus, and retina in the anesthetized cat
- PMID: 9698331
- PMCID: PMC6793201
- DOI: 10.1523/JNEUROSCI.18-16-06395.1998
Synchronization of visual responses between the cortex, lateral geniculate nucleus, and retina in the anesthetized cat
Abstract
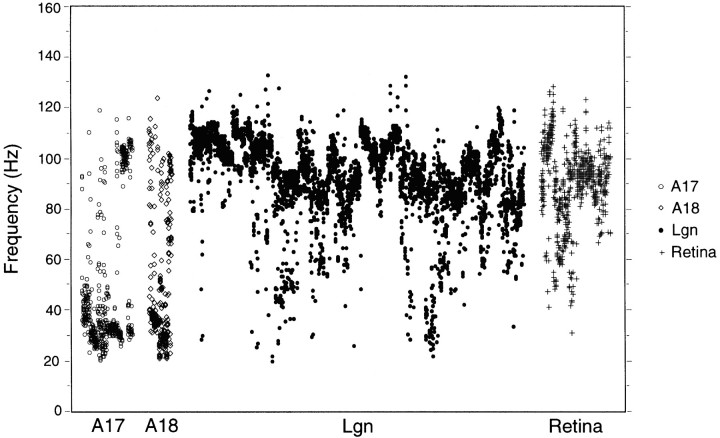
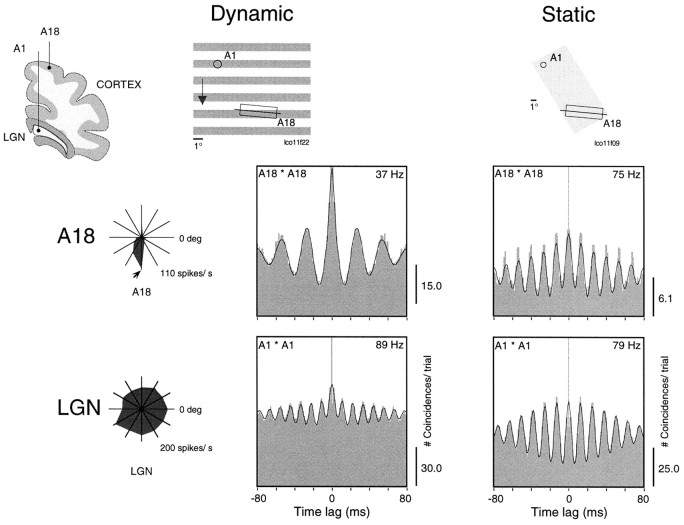
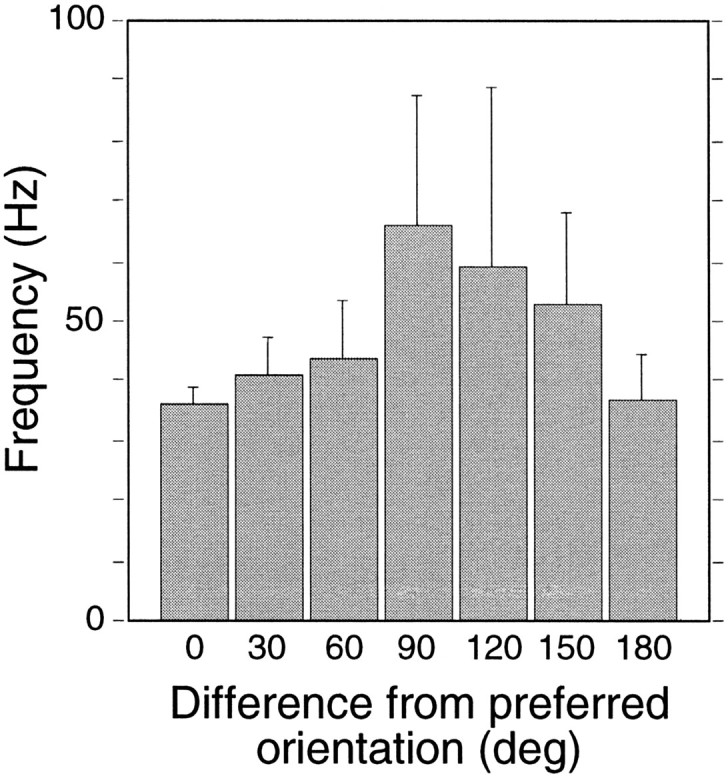
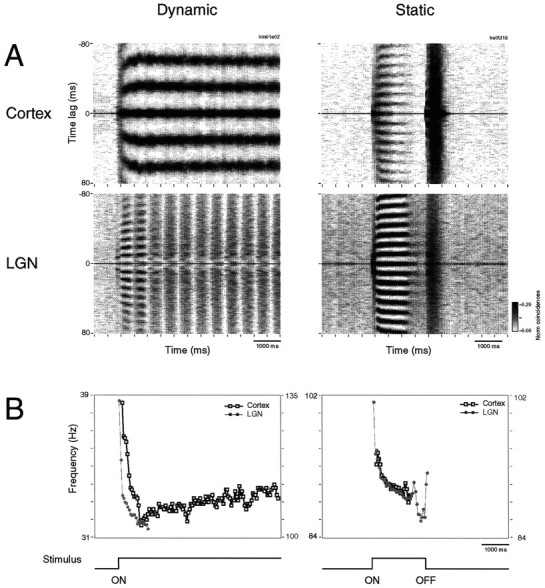
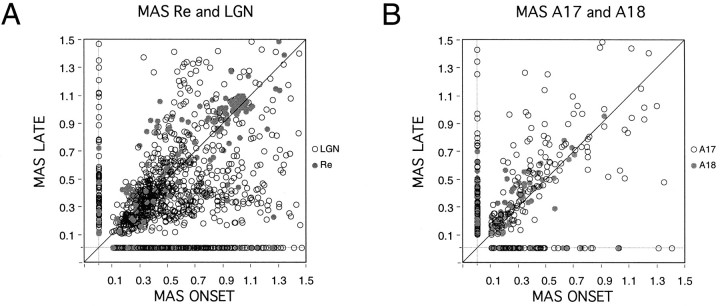
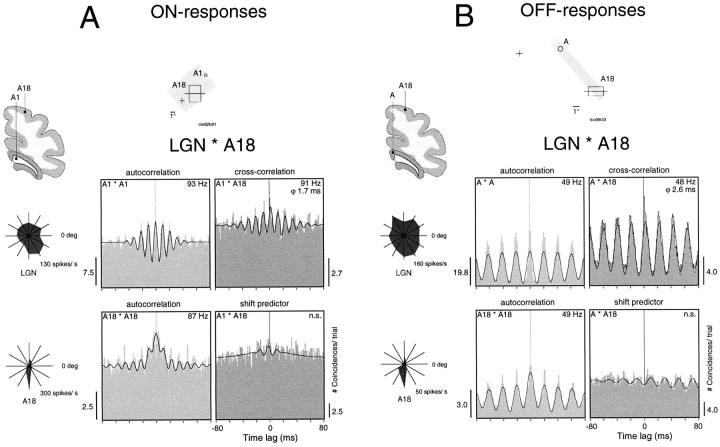
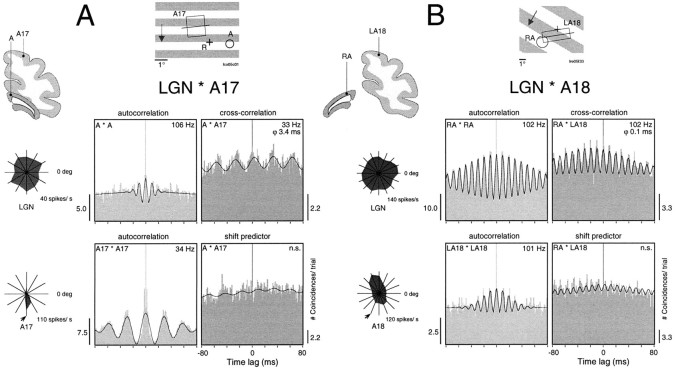
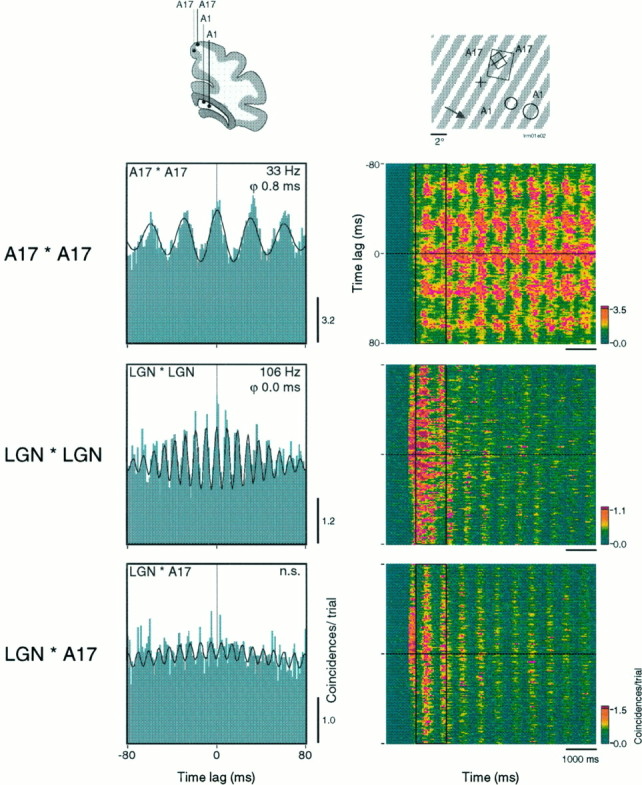
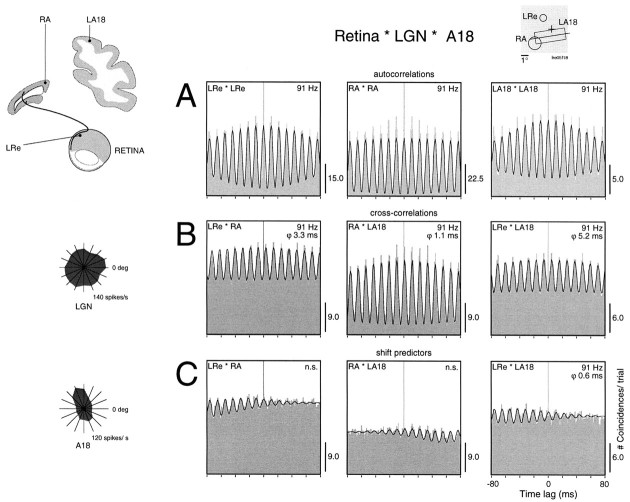
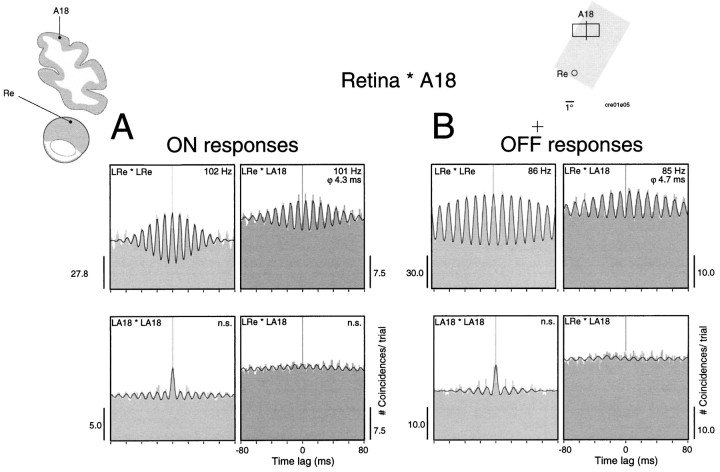
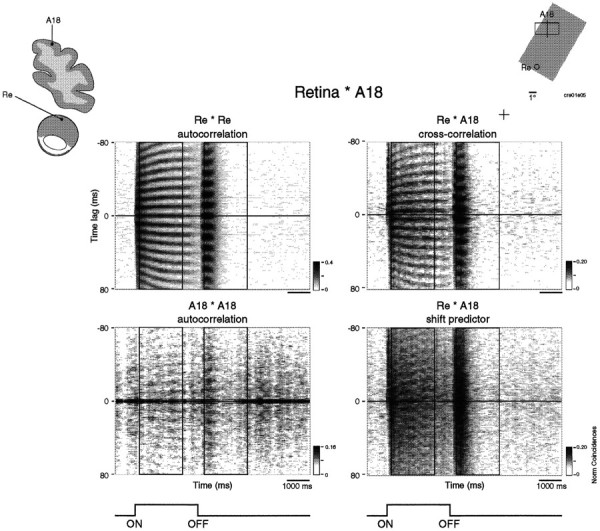
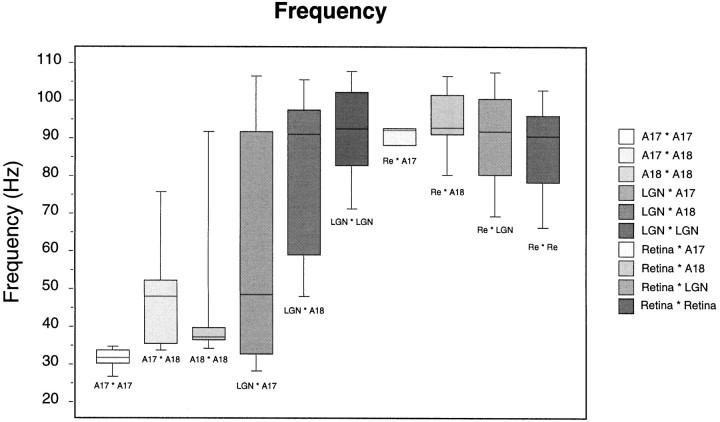
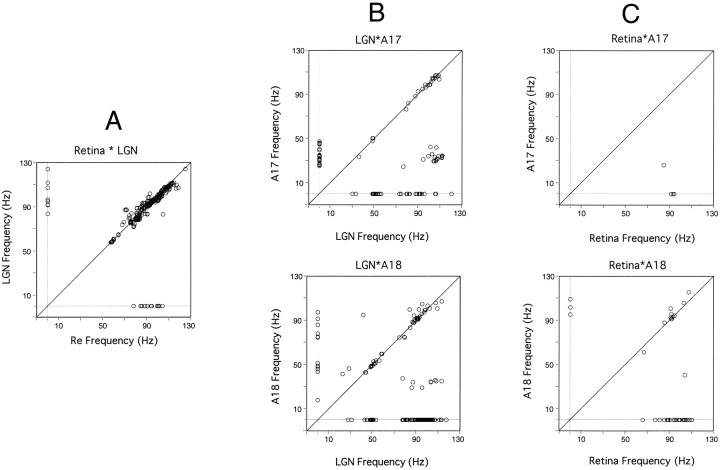
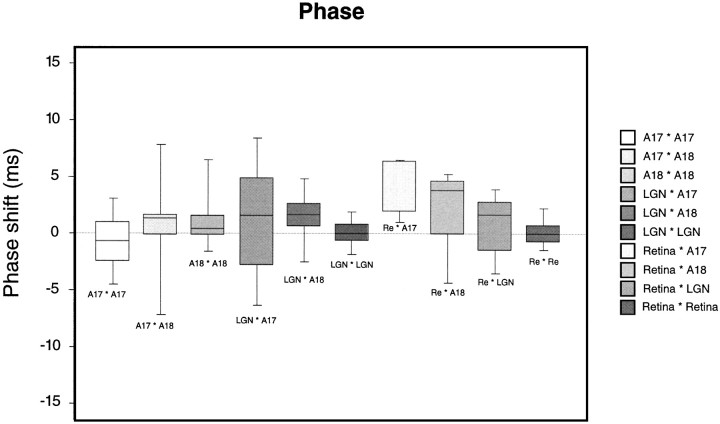
Synchronization of spatially distributed responses in the cortex is often associated with periodic activity. Recently, synchronous oscillatory patterning was described for visual responses in retinal ganglion cells that is reliably transmitted by the lateral geniculate nucleus (LGN), raising the question of whether oscillatory inputs contribute to synchronous oscillatory responses in the cortex. We have made simultaneous multi-unit recordings from visual areas 17 and 18 as well as the LGN and the retina to examine the interactions between subcortical and cortical synchronization mechanisms. Strong correlations of oscillatory responses were observed between retina, LGN, and cortex, indicating that cortical neurons can become synchronized by oscillatory activity relayed through the LGN. This feedforward synchronization occurred with oscillation frequencies in the range of 60-120 Hz and was most pronounced for responses to stationary flashed stimuli and more frequent for cells in area 18 than in area 17. In response to moving stimuli, by contrast, subcortical and cortical oscillations dissociated, proving the existence of independent subcortical and cortical mechanisms. Subcortical oscillations maintained their high frequencies but became transient. Cortical oscillations were now dominated by a cortical synchronizing mechanism operating in the 30-60 Hz frequency range. When the cortical mechanism dominated, LGN responses could become phase-locked to the cortical oscillations via corticothalamic feedback. In summary, synchronization of cortical responses can result from two independent but interacting mechanisms. First, a transient feedforward synchronization to high-frequency retinal oscillations, and second, an intracortical mechanism, which operates in a lower frequency range and induces more sustained synchronization.
Figures
References
-
- Abeles M. Corticonics. Neural circuits of the cerebral cortex. Cambridge UP; Cambridge: 1991.
-
- Alonso J-M, Usrey WM, Reid RC. Precisely correlated firing in cells of the lateral geniculate nucleus. Nature. 1996;383:815–819. - PubMed
-
- Arnett DW. Correlation analysis of units recorded in the cat dorsal lateral geniculate nucleus. Exp Brain Res. 1975;24:111–130. - PubMed
-
- Blake R, Camisa R. Temporal aspects of spatial vision in cats. Exp Brain Res. 1977;28:325–333. - PubMed
-
- Cleland BG, Dubin MW, Levick WR. Simultaneous recording of input and output of lateral geniculate neurons. Nature. 1971a;231:191–192. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous