

Evidence that trigeminal brainstem interneurons form subpopulations to produce different forms of mastication in the rabbit
- PMID: 9698335
- PMCID: PMC6793206
- DOI: 10.1523/JNEUROSCI.18-16-06466.1998
Evidence that trigeminal brainstem interneurons form subpopulations to produce different forms of mastication in the rabbit
Abstract
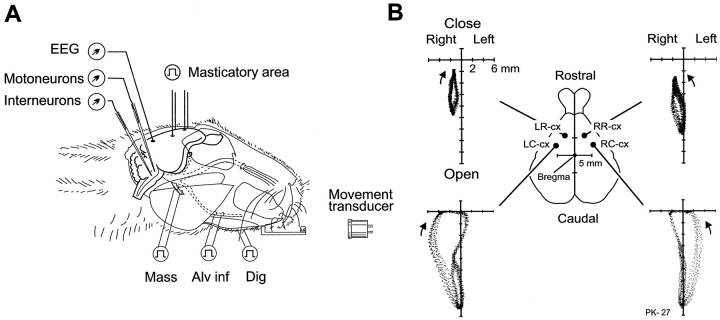
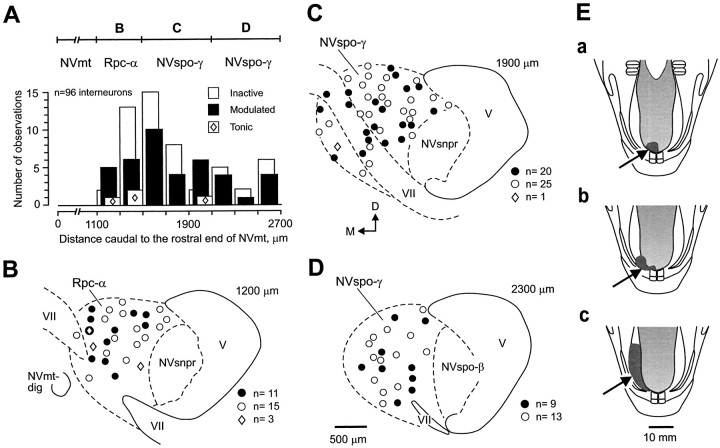
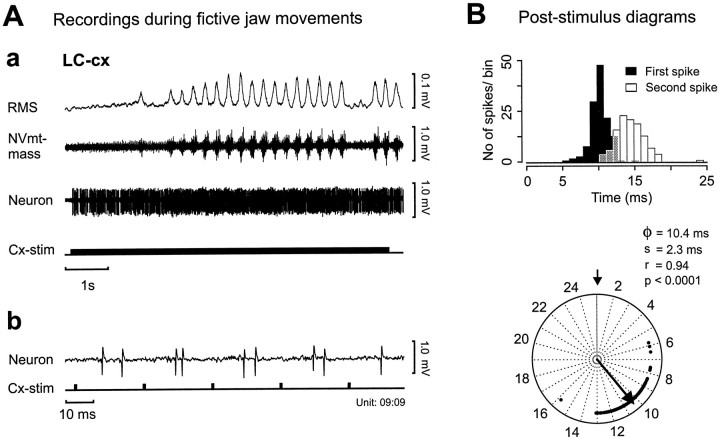
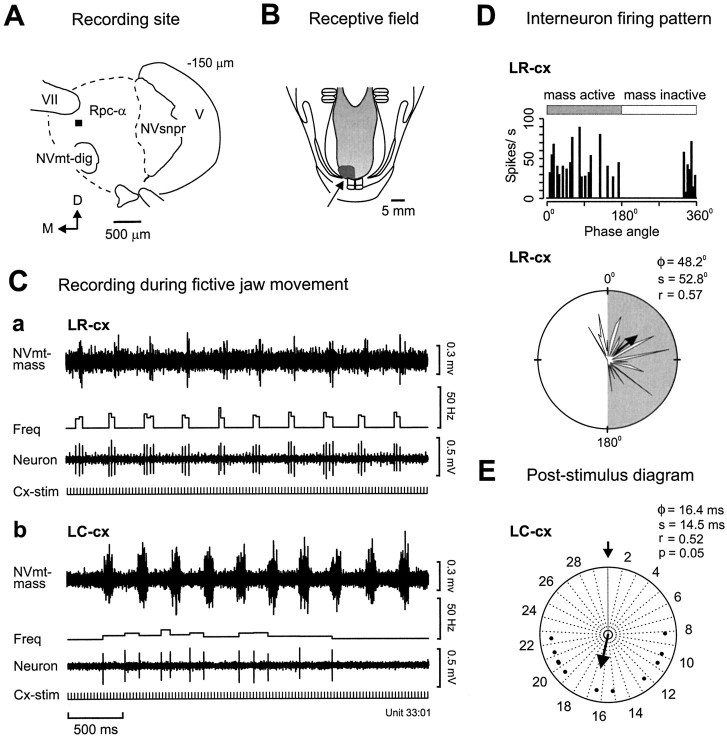
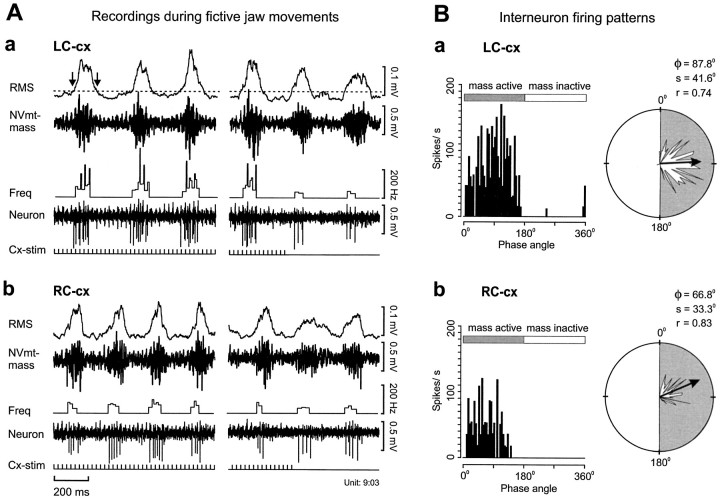
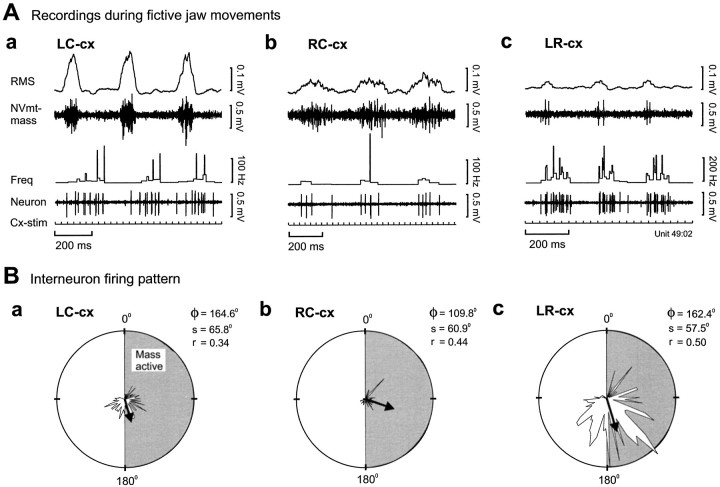
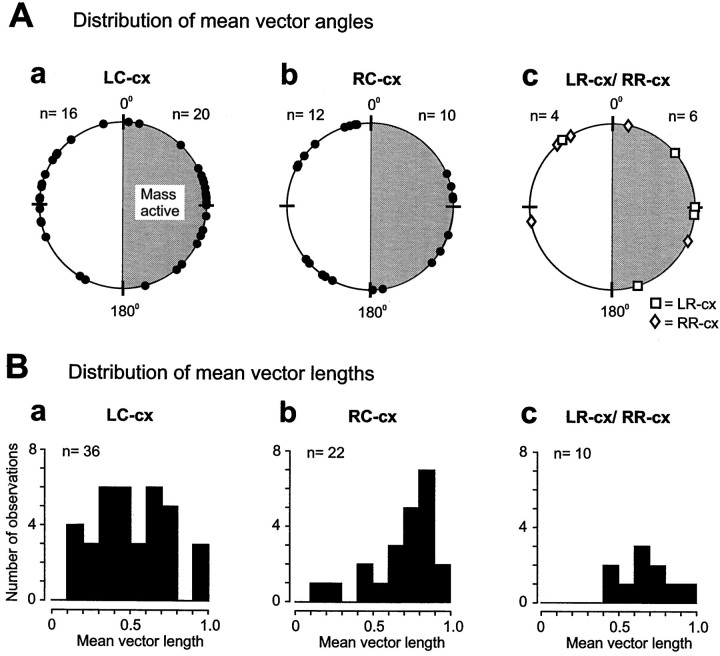
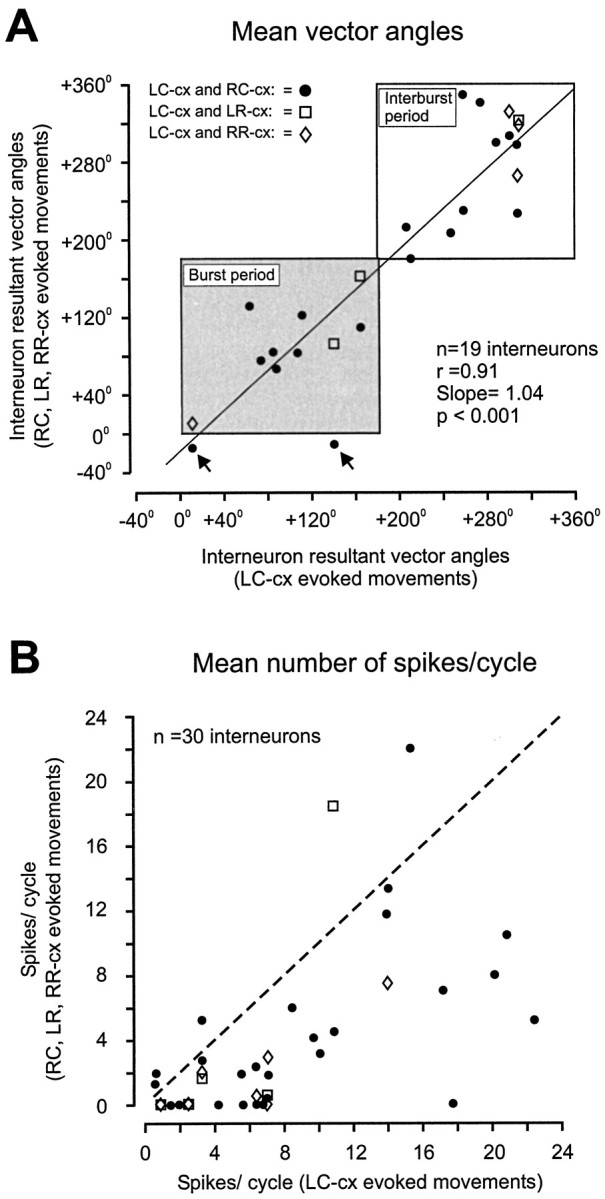
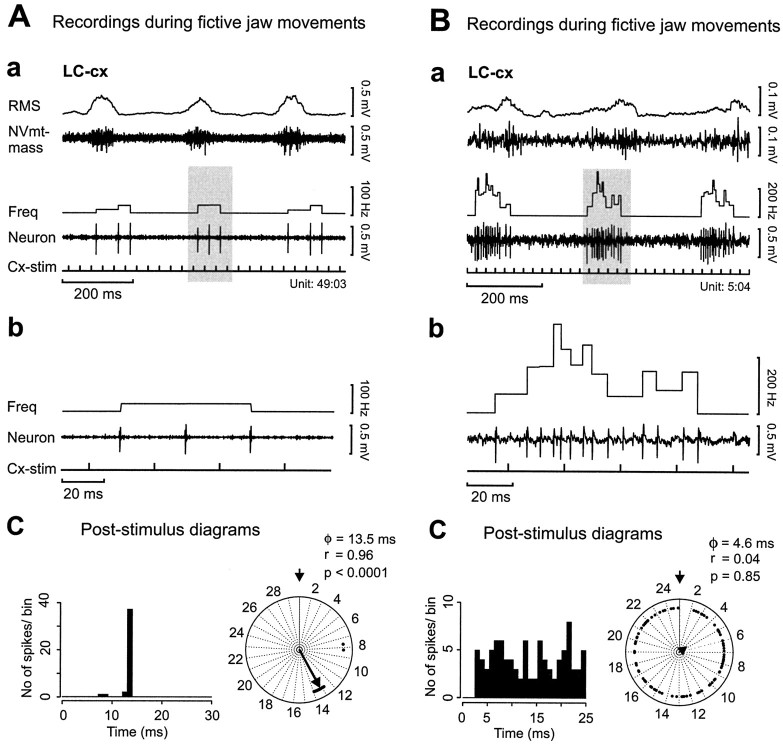
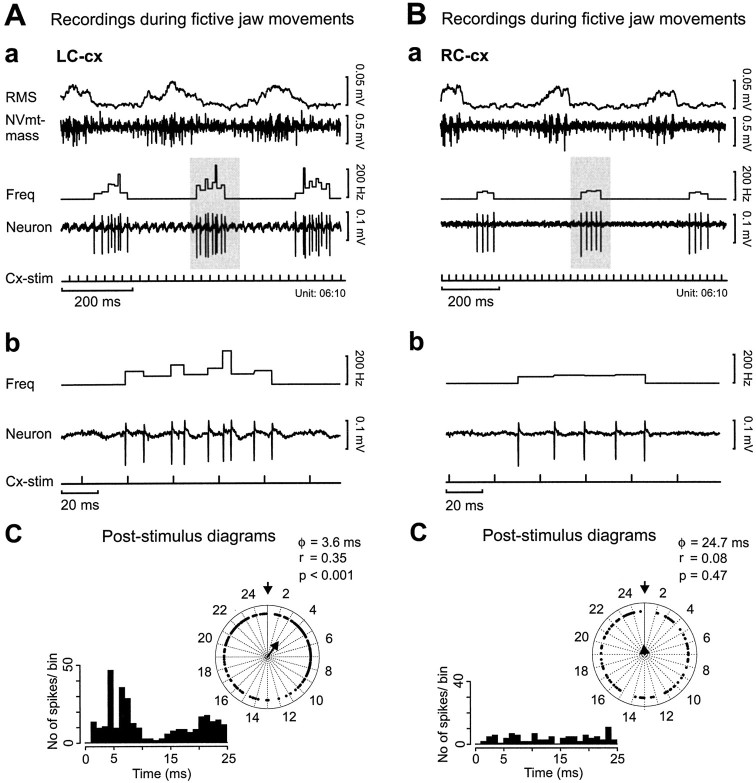
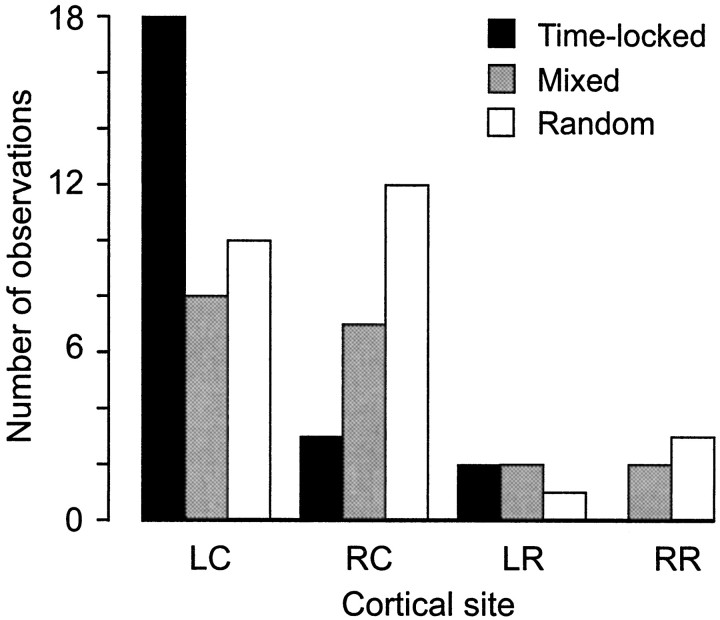
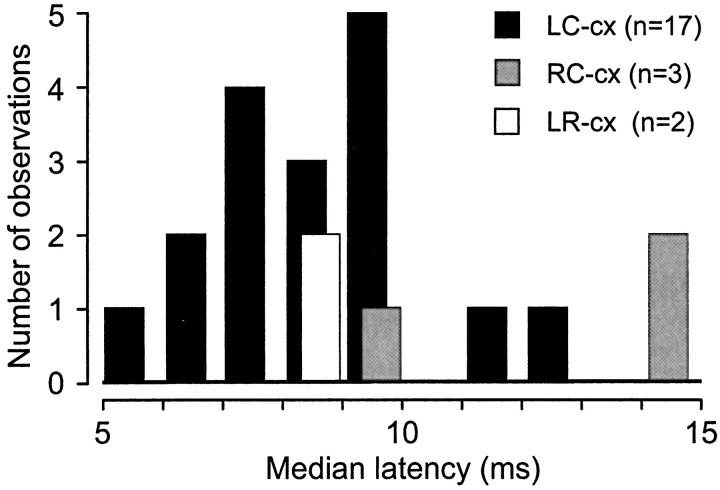
To determine how trigeminal brainstem interneurons pattern different forms of rhythmical jaw movements, four types of motor patterns were induced by electrical stimulation within the cortical masticatory areas of rabbits. After these were recorded, animals were paralyzed and fictive motor output was recorded with an extracellular microelectrode in the trigeminal motor nucleus. A second electrode was used to record from interneurons within the lateral part of the parvocellular reticular formation (Rpc-alpha, n = 28) and gamma- subnucleus of the oral nucleus of the spinal trigeminal tract (NVspo-gamma, n = 68). Both of these areas contain many interneurons projecting to the trigeminal motor nucleus. The basic characteristics of the four movement types evoked before paralysis were similar to those seen after the neuromuscular blockade, although cycle duration was significantly decreased for all patterns. Interneurons showed three types of firing pattern: 54% were inactive, 42% were rhythmically active, and 4% had a tonic firing pattern. Neurons within the first two categories were intermingled in Rpc-alpha and NVspo-gamma: 48% of rhythmic neurons were active during one movement type, 35% were active during two, and 13% were active during three or four patterns. Most units fired during either the middle of the masseter burst or interburst phases during fictive movements evoked from the left caudal cortex. In contrast, there were no tendencies toward a preferred coupling of interneuron activity to any particular phase of the cycle during stimulation of other cortical sites. It was concluded that the premotoneurons that form the final commands to trigeminal motoneurons are organized into subpopulations according to movement pattern.
Figures
References
-
- Appenteng K, Girdlestone D. Transneuronal transport of wheat-germ agglutinin-conjugated horseradish peroxidase into trigeminal interneurones of the cat. J Comp Neurol. 1987;258:387–393. - PubMed
-
- Appenteng K, Conyers L, Curtis J, Moore J. Monosynaptic connections of single V interneurones to the contralateral V motor nucleus in anaesthetized rats. Brain Res. 1990;514:128–130. - PubMed
-
- Batschelet E. Circular statistics in biology. Academic; New York: 1981.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources