

Cell-specific transcription of the smooth muscle gamma-actin gene requires both positive- and negative-acting cis elements
- PMID: 9699483
- PMCID: PMC6190202
Cell-specific transcription of the smooth muscle gamma-actin gene requires both positive- and negative-acting cis elements
Abstract
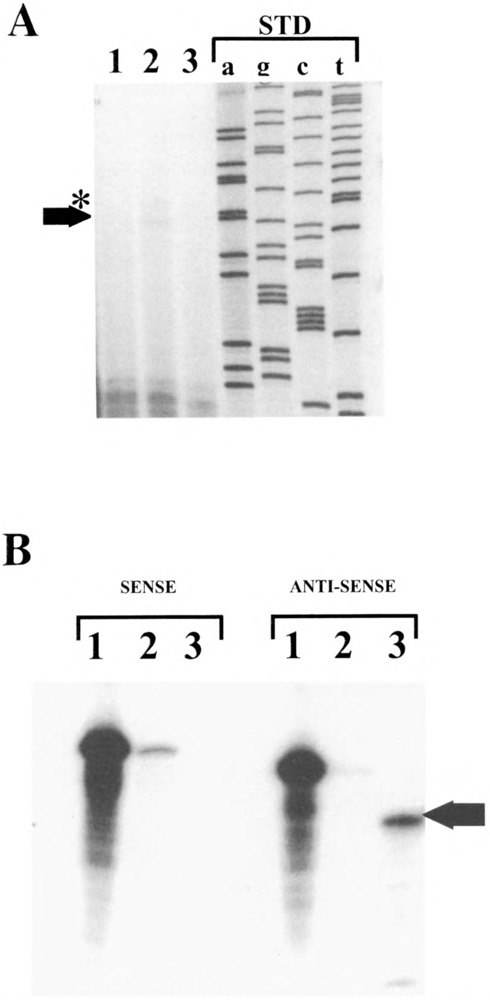
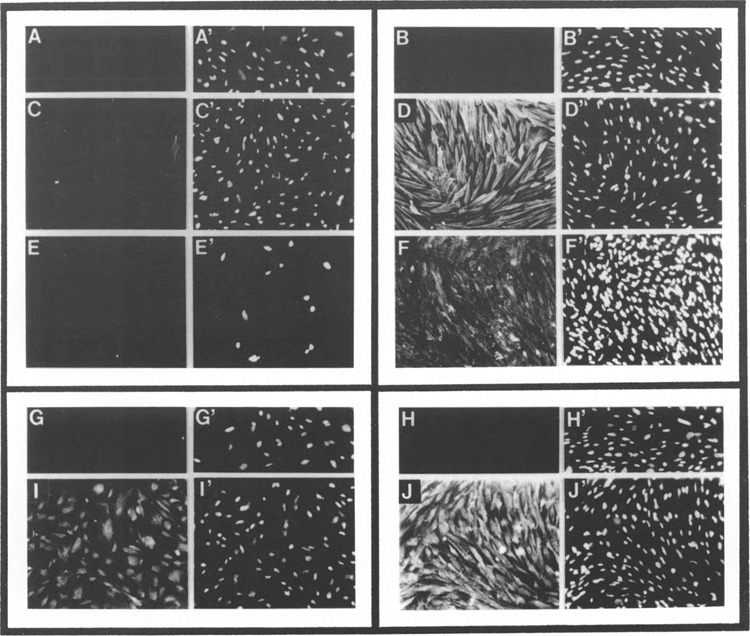
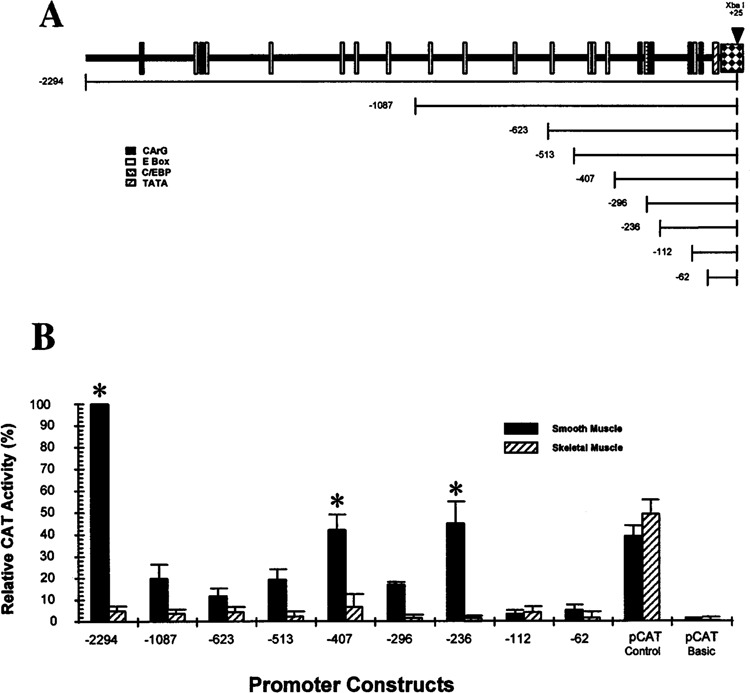
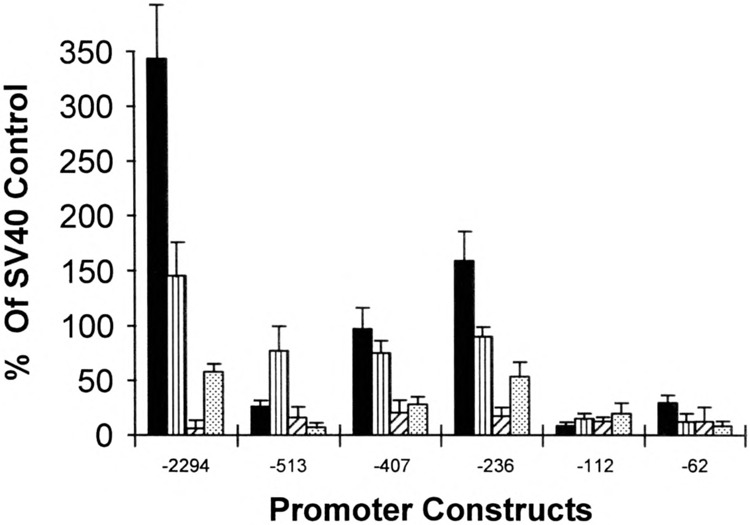
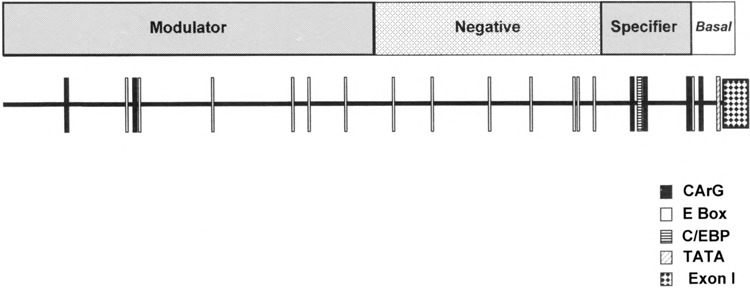
We have characterized the function of putative regulatory sequences upon the smooth muscle transcription of the SMGA gene, using promoter deletion analyses. We demonstrate that the SMGA promoter contains four domains: a basal promoter (-1 to -100), a smooth muscle specifier sequence (-100 to -400), a negative regulator (-400 to -1000), and a smooth muscle-specific modulator (-1000 to -2000). The basal or core promoter supports equivalent transcription in both smooth and skeletal muscle cells. Addition of sequences containing a CArG motif juxtaposed to an E-box element stimulates smooth muscle transcription by five- to sixfold compared to skeletal muscle. This smooth muscle-specific segment is maintained for about 200 bp, after which is a segment of DNA that appears to inhibit the transcriptional capacity of the SMGA promoter in smooth muscle cells. Within the boundary between the smooth muscle specifier and negative regulatory sequences (-400 to -500) are three E-box elements. The smooth muscle modulator domain contains two CArG elements and multiple E-boxes. When added to the SMGA promoter it causes an additional three- to fivefold increase in smooth muscle-specific transcription over that stimulated by the smooth muscle specifier domain. Thus, our studies show that the appropriate cell-specific transcription of the SMGA gene involves complex interactions directed by multiple cis-acting elements. Moreover, our characterization of a cell culture system employing embryonic gizzard smooth muscle cells lays the foundation for further molecular analyses of factors that regulate or control SMGA and other smooth muscle genes during differentiation.
Figures

Similar articles
-
The developmentally regulated expression of serum response factor plays a key role in the control of smooth muscle-specific genes.Dev Biol. 1998 Feb 1;194(1):18-37. doi: 10.1006/dbio.1997.8808. Dev Biol. 1998. PMID: 9473329
-
The smooth muscle gamma-actin gene promoter is a molecular target for the mouse bagpipe homologue, mNkx3-1, and serum response factor.J Biol Chem. 2000 Dec 15;275(50):39061-72. doi: 10.1074/jbc.M006532200. J Biol Chem. 2000. PMID: 10993896
-
Smooth muscle gamma-actin promoter regulation by RhoA and serum response factor signaling.Biochim Biophys Acta. 2003 Jul 28;1628(2):133-9. doi: 10.1016/s0167-4781(03)00122-2. Biochim Biophys Acta. 2003. PMID: 12890560
-
Differentiating one smooth operator from another.J Mol Med (Berl). 1999 Feb;77(2):255-7. doi: 10.1007/s001090050347. J Mol Med (Berl). 1999. PMID: 10023778 Review. No abstract available.
-
Developmental regulation of elastin gene expression.Crit Rev Eukaryot Gene Expr. 1997;7(1-2):1-10. doi: 10.1615/critreveukargeneexpr.v7.i1-2.10. Crit Rev Eukaryot Gene Expr. 1997. PMID: 9034712 Review.
Cited by
-
The smooth muscle gamma-actin gene is androgen responsive in prostate epithelia.Gene Expr. 2002;10(5-6):201-11. doi: 10.3727/000000002783992424. Gene Expr. 2002. PMID: 12450213 Free PMC article.
-
Transforming growth factor-beta induction of smooth muscle cell phenotpye requires transcriptional and post-transcriptional control of serum response factor.J Biol Chem. 2002 Feb 22;277(8):6287-95. doi: 10.1074/jbc.M106649200. Epub 2001 Dec 11. J Biol Chem. 2002. PMID: 11741973 Free PMC article.
-
Intracellular trafficking of nucleic acids.Expert Opin Drug Deliv. 2004 Nov;1(1):127-40. doi: 10.1517/17425247.1.1.127. Expert Opin Drug Deliv. 2004. PMID: 16296725 Free PMC article. Review.
-
Serum response factor is crucial for actin cytoskeletal organization and focal adhesion assembly in embryonic stem cells.J Cell Biol. 2002 Feb 18;156(4):737-50. doi: 10.1083/jcb.200106008. Epub 2002 Feb 11. J Cell Biol. 2002. PMID: 11839767 Free PMC article.
-
Intracellular trafficking of plasmids for gene therapy: mechanisms of cytoplasmic movement and nuclear import.Curr Gene Ther. 2006 Dec;6(6):671-681. doi: 10.2174/156652306779010688. Curr Gene Ther. 2006. PMID: 17168698 Free PMC article. Review.
References
-
- Al-Molish M. I.; Dubes G. R. Enhanced DNA transfer with dextrans. J. Gen. Virol. 18:189–193; 1973. - PubMed
-
- Balczon R.; West K. The identification of mammalian centrosomal antigens using human autoimmune anti-centrosome antisera. Cell Motil. Cytoskeleton 20:121–135; 1991. - PubMed
-
- Blank R. S.; McQuinn T. C.; Yin K. C.; Thompson M. M.; Takeyasu K.; Schwartz R. J.; Owens G. K. Elements of the smooth muscle α-actin promoter required in cis for transcriptional activation in smooth muscle. J. Biol. Chem. 267:984–989; 1992. - PubMed
-
- Breathnach R.; Chambon P. Organization and expression of eukaryotic split genes coding for proteins. Annu. Rev. Biochem. 50:349–383; 1981. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources