

Mitotic centromere-associated kinesin is important for anaphase chromosome segregation
- PMID: 9700166
- PMCID: PMC2148171
- DOI: 10.1083/jcb.142.3.787
Mitotic centromere-associated kinesin is important for anaphase chromosome segregation
Abstract
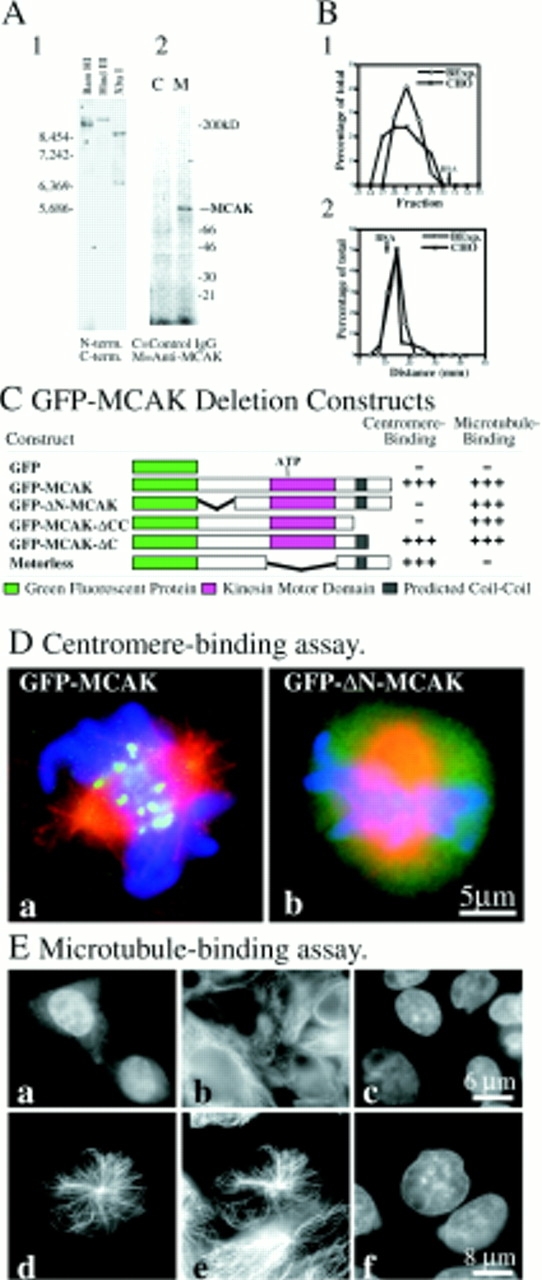
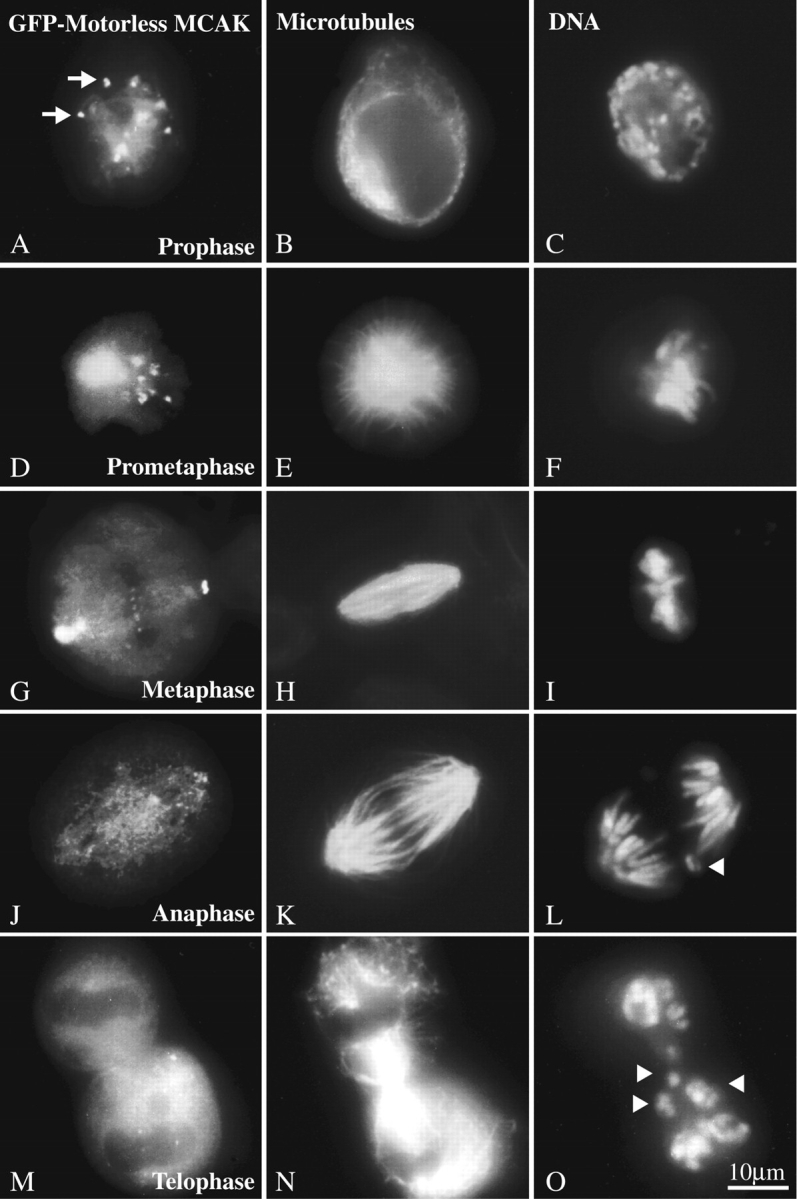
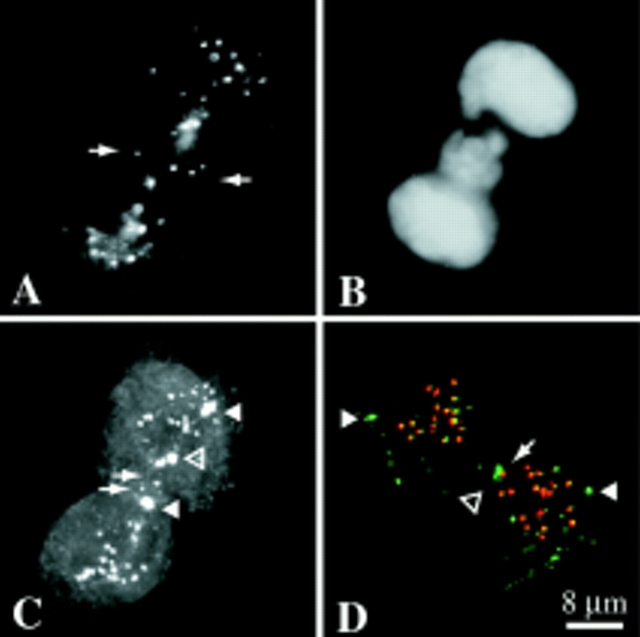
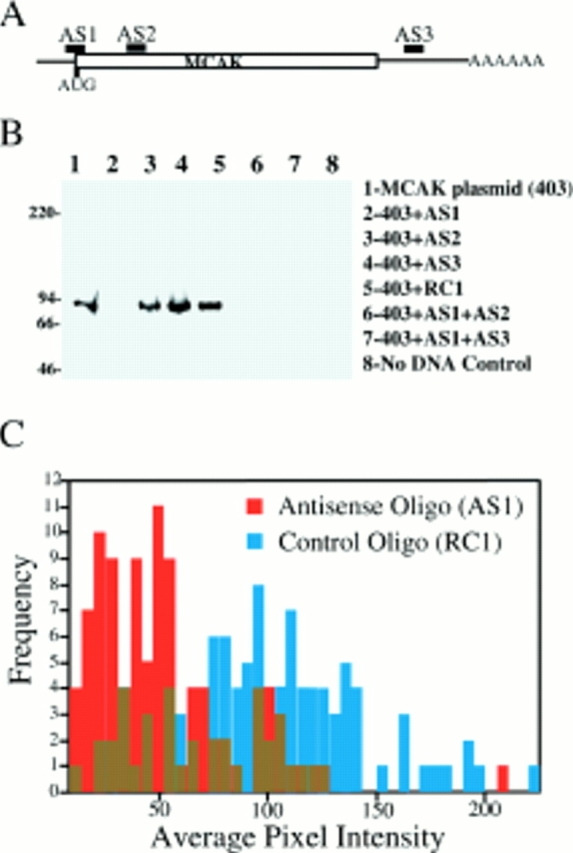
Mitotic centromere-associated kinesin (MCAK) is recruited to the centromere at prophase and remains centromere associated until after telophase. MCAK is a homodimer that is encoded by a single gene and has no associated subunits. A motorless version of MCAK that binds centromeres but not microtubules disrupts chromosome segregation during anaphase. Antisense-induced depletion of MCAK results in the same defect. MCAK overexpression induces centromere-independent bundling and eventual loss of spindle microtubule polymer suggesting that centromere-associated bundling and/or depolymerization activity is required for anaphase. Live cell imaging indicates that MCAK may be required to coordinate the onset of sister centromere separation.
Figures
References
-
- Ault JG, DeMarco AJ, Salmon ED, Rieder CL. Studies on the ejection properties of asters: astral microtubule turnover influences the oscillatory behavior and positioning of mono-oriented chromosomes. J Cell Sci. 1991;99:701–710. - PubMed
-
- Bloom K. The centromere frontier: Kinetochore components, microtubule-based motility, and the CEN-value paradox. Cell. 1993;73:621–624. - PubMed
-
- Brown KD, Wood KW, Cleveland DW. The kinesin-like protein CENP-E is kinetochore-associated throughout poleward chromosome segregation during anaphase-A. J Cell Sci. 1996;109:961–969. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
