

The fibronectin domain ED-A is crucial for myofibroblastic phenotype induction by transforming growth factor-beta1
- PMID: 9700173
- PMCID: PMC2148176
- DOI: 10.1083/jcb.142.3.873
The fibronectin domain ED-A is crucial for myofibroblastic phenotype induction by transforming growth factor-beta1
Abstract
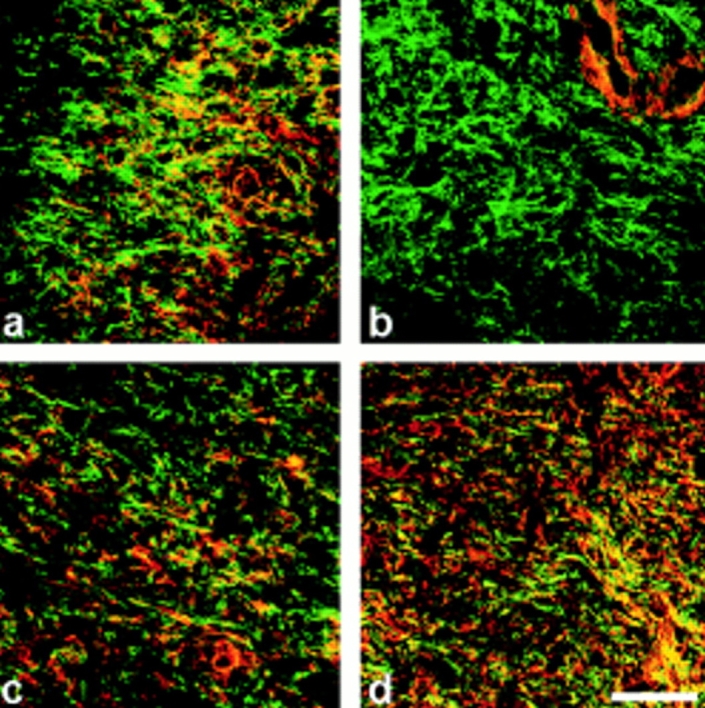
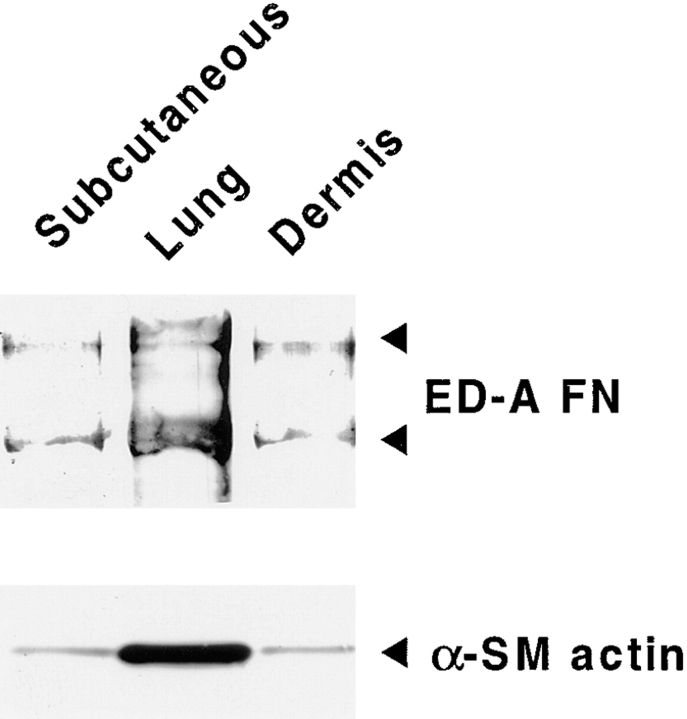
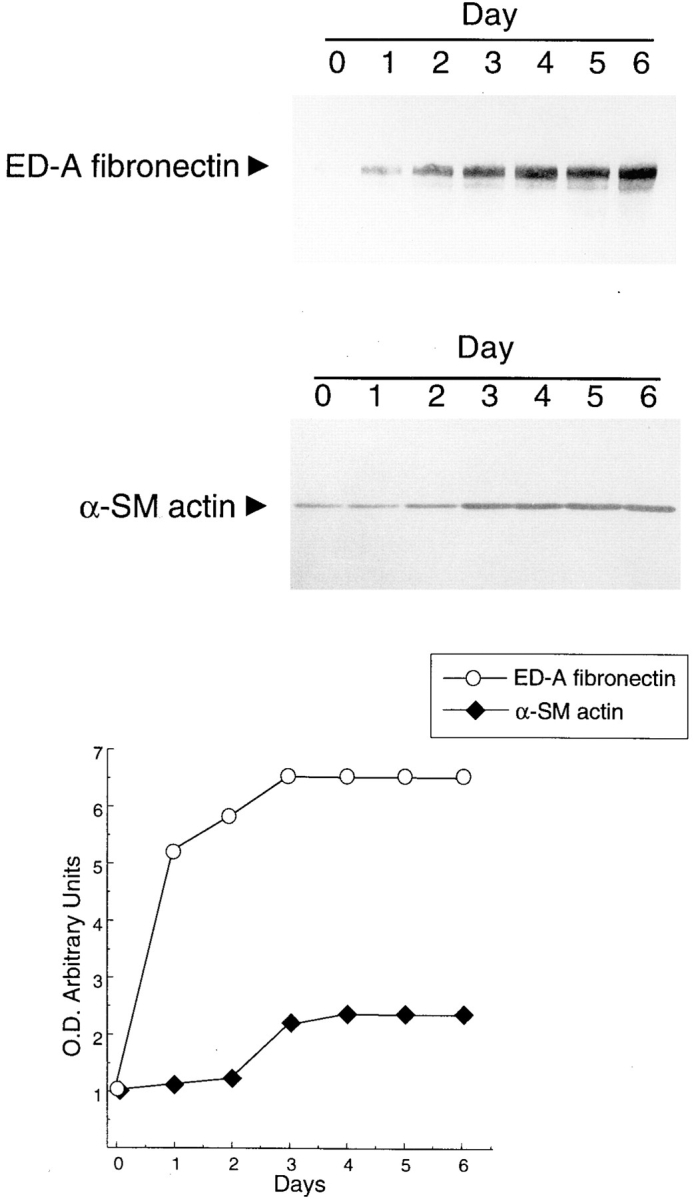
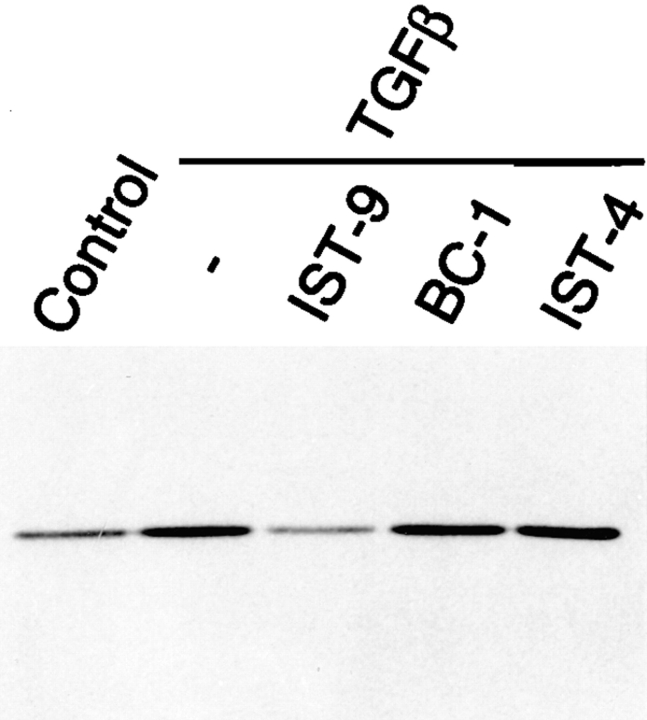
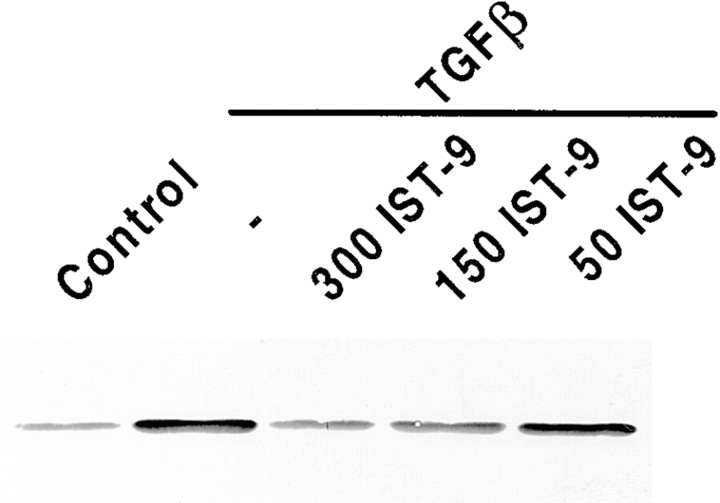
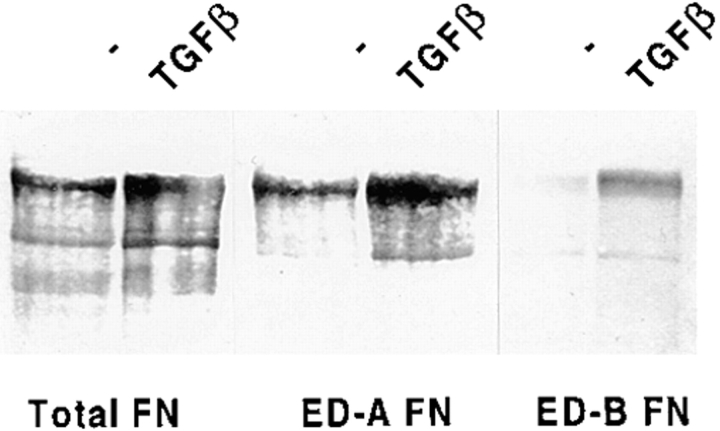
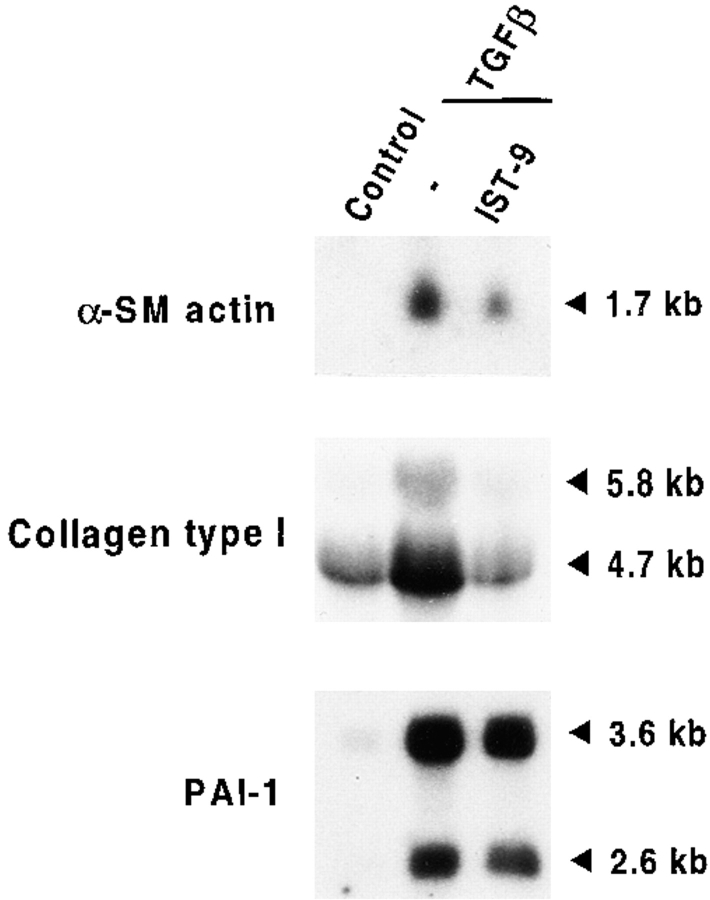
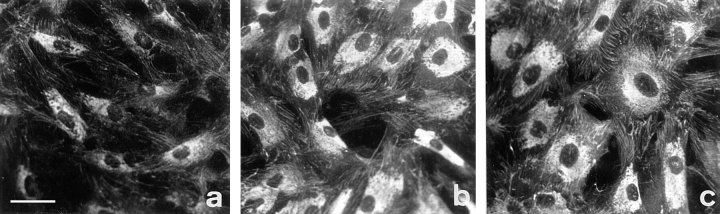
Transforming growth factor-beta1 (TGFbeta1), a major promoter of myofibroblast differentiation, induces alpha-smooth muscle (sn) actin, modulates the expression of adhesive receptors, and enhances the synthesis of extracellular matrix (ECM) molecules including ED-A fibronectin (FN), an isoform de novo expressed during wound healing and fibrotic changes. We report here that ED-A FN deposition precedes alpha-SM actin expression by fibroblasts during granulation tissue evolution in vivo and after TGFbeta1 stimulation in vitro. Moreover, there is a correlation between in vitro expression of alpha-SM actin and ED-A FN in different fibroblastic populations. Seeding fibroblasts on ED-A FN does not elicit per se alpha-SM actin expression; however, incubation of fibroblasts with the anti-ED-A monoclonal antibody IST-9 specifically blocks the TGFbeta1-triggered enhancement of alpha-SM actin and collagen type I, but not that of plasminogen activator inhibitor-1 mRNA. Interestingly, the same inhibiting action is exerted by the soluble recombinant domain ED-A, but neither of these inhibitory agents alter FN matrix assembly. Our findings indicate that ED-A-containing polymerized FN is necessary for the induction of the myofibroblastic phenotype by TGFbeta1 and identify a hitherto unknown mechanism of cytokine-determined gene stimulation based on the generation of an ECM-derived permissive outside in signaling, under the control of the cytokine itself.
Figures

References
-
- Balza E, Borsi L, Allemmanni G, Zardi L. Transforming Growth Factor β regulates the levels of different fibronectin isoforms in normal cultured fibroblasts. FEBS (Fed Eur Biochem Soc) Lett. 1988;228:42–44. - PubMed
-
- Berndt A, Kosmehl H, Mandel U, Gabler U, Luo X, Celada D, Zardi L, Katenkamp D. TGFβ and bFGF synthesis and localization in Dupuytren's disease (nodular palmar fibrosis) relative to cellular activity, myofibroblast phenotype and oncofetal variants of fibronectin. Histochemical J. 1995;27:1014–1020. - PubMed
-
- Bochaton-Piallat ML, Gabbiani G, Pepper MS. Plasminogen activator expression in rat arterial smooth muscle cells depends on their phenotype and is modulated by cytokines. Circ Res. 1998;82:1086–1093. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
