

Induction of a carbon-starvation-related proteolysis in whole maize plants submitted to Light/Dark cycles and to extended darkness
- PMID: 9701583
- PMCID: PMC34891
- DOI: 10.1104/pp.117.4.1281
Induction of a carbon-starvation-related proteolysis in whole maize plants submitted to Light/Dark cycles and to extended darkness
Abstract
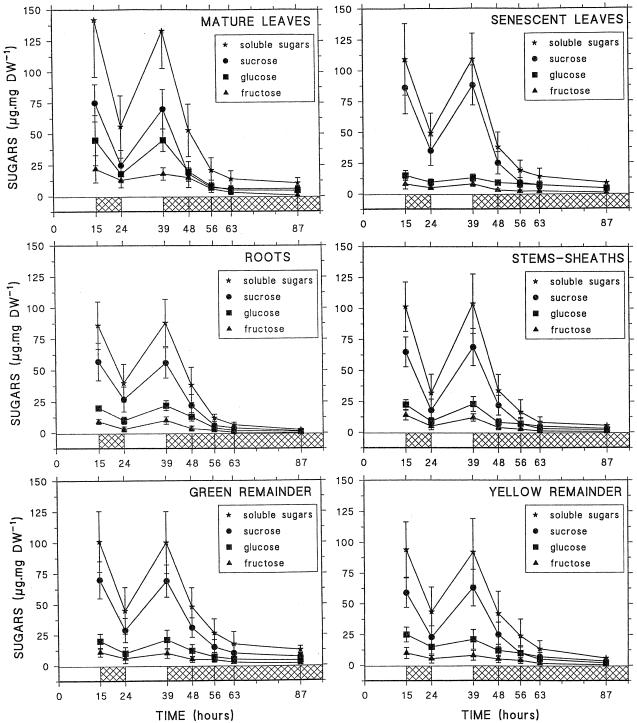
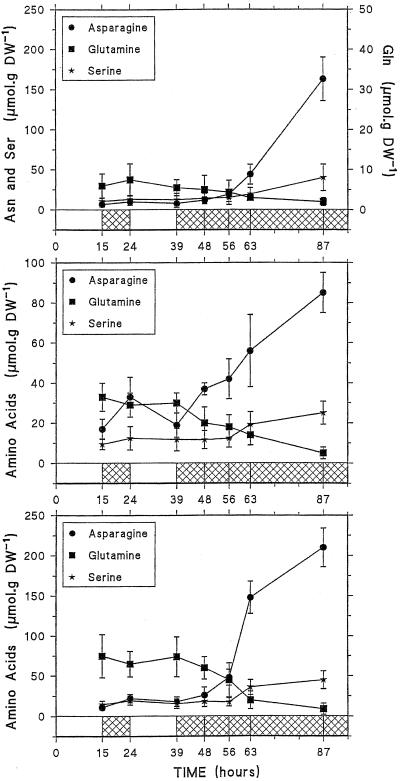
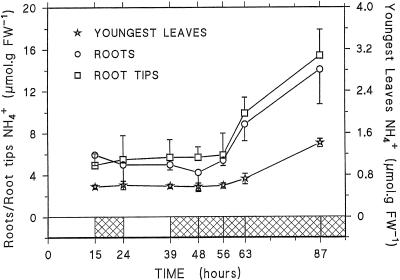
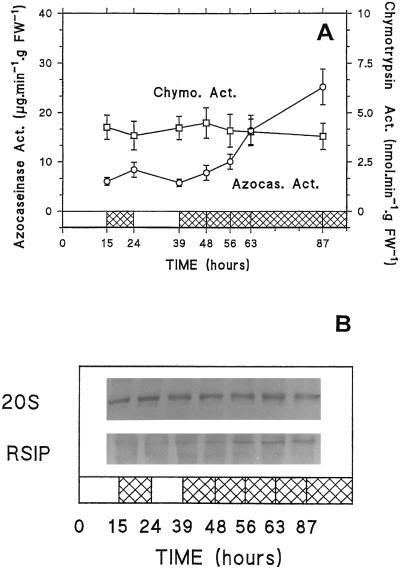
Three-week-old maize (Zea mays L.) plants were submitted to light/dark cycles and to prolonged darkness to investigate the occurrence of sugar-limitation effects in different parts of the whole plant. Soluble sugars fluctuated with light/dark cycles and dropped sharply during extended darkness. Significant decreases in protein level were observed after prolonged darkness in mature roots, root tips, and young leaves. Glutamine and asparagine (Asn) changed in opposite ways, with Asn increasing in the dark. After prolonged darkness the increase in Asn accounted for most of the nitrogen released by protein breakdown. Using polyclonal antibodies against a vacuolar root protease previously described (F. James, R. Brouquisse, C. Suire, A. Pradet, P. Raymond [1996] Biochem J 320: 283-292) or the 20S proteasome, we showed that the increase in proteolytic activities was related to an enrichment of roots in the vacuolar protease, with no change in the amount of 20S proteasome in either roots or leaves. Our results show that no significant net proteolysis is induced in any part of the plant during normal light/dark cycles, although changes in metabolism and growth appear soon after the beginning of the dark period, and starvation-related proteolysis probably appears in prolonged darkness earlier in sink than in mature tissues.
Figures


References
-
- Amthor JS, McCree KJ (1990) Carbon balance of stressed plants: a conceptual model for integrating research results. In RG Alscher, JC Cumming, eds, Stress Responses in Plants: Adaptation and Acclimation Mechanisms. Wiley-Liss & Sons, Inc., New York, pp 1–15
-
- Aubert S, Gout E, Bligny R, Marty-Mazars D, Barrieu F, Alabouvette J, Marty F, Douce R. Ultrastructural and biochemical characterization of autophagy in higher plant cells subjected to carbon deprivation: control by the supply of mitochondria with respiratory substrates. J Cell Biol. 1996;133:1251–1263. - PMC - PubMed
-
- Bingham IJ, Stevenson EA. Control of root growth: effects of carbohydrates on the extension, branching and rate of respiration of different fractions of wheat roots. Physiol Plant. 1994;88:149–158.
LinkOut - more resources
Full Text Sources
