Differentiation and cancer in the mammary gland: shedding light on an old dichotomy
- PMID: 9709809
- PMCID: PMC3867313
- DOI: 10.1016/s0065-230x(08)60741-1
Differentiation and cancer in the mammary gland: shedding light on an old dichotomy
Abstract
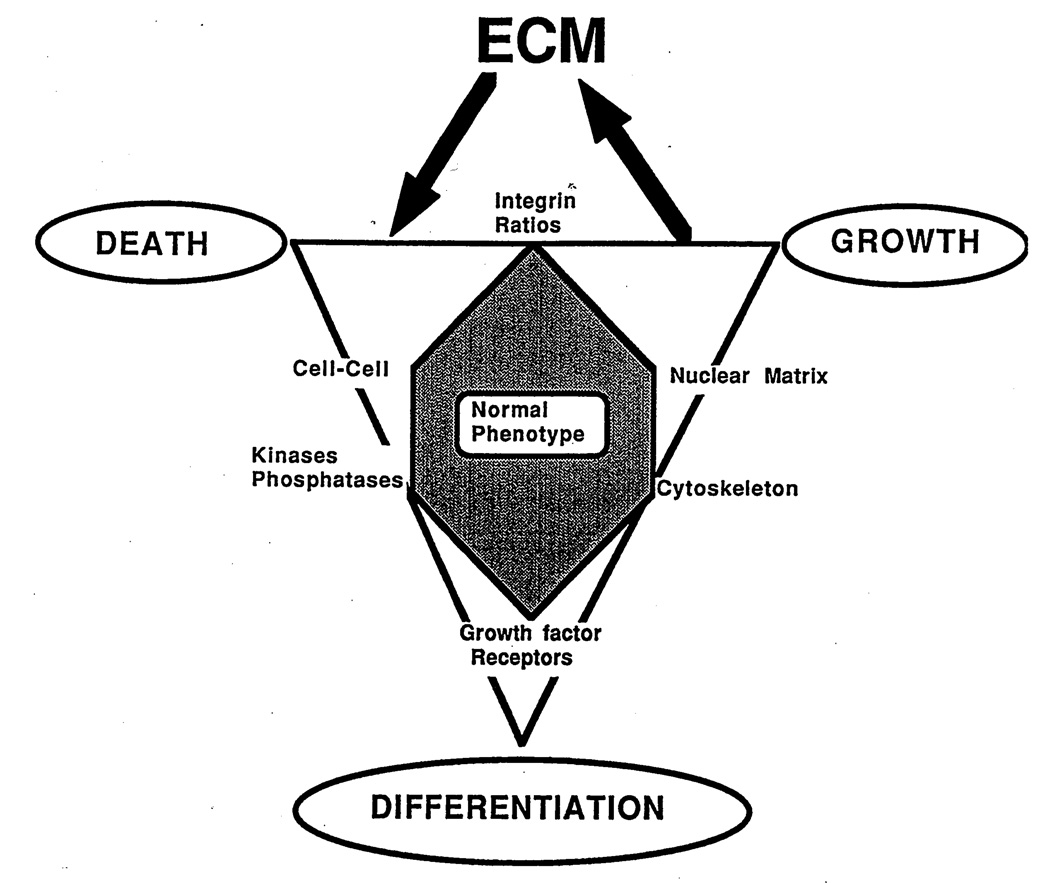
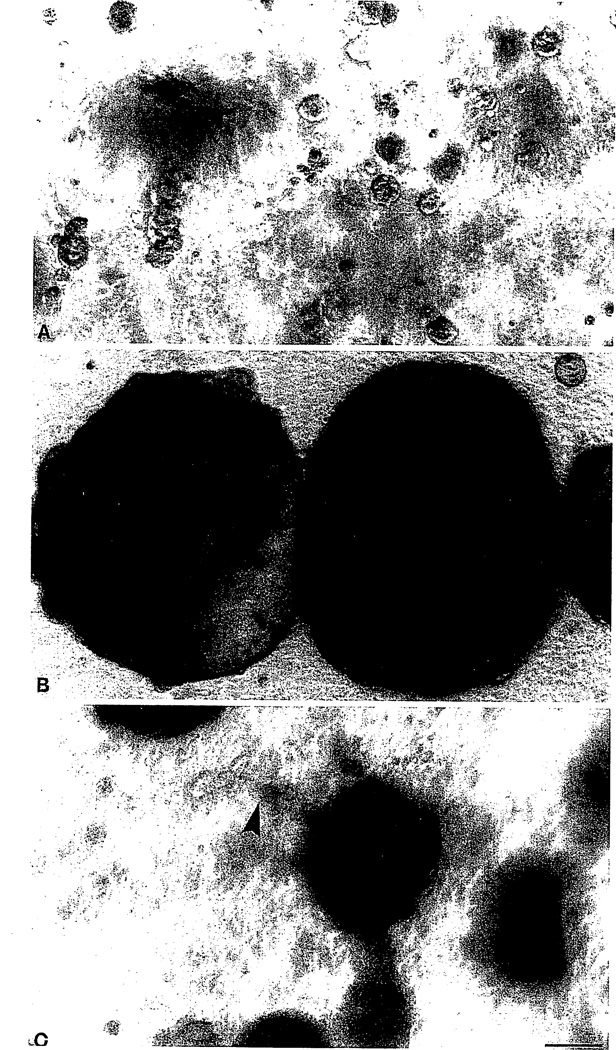
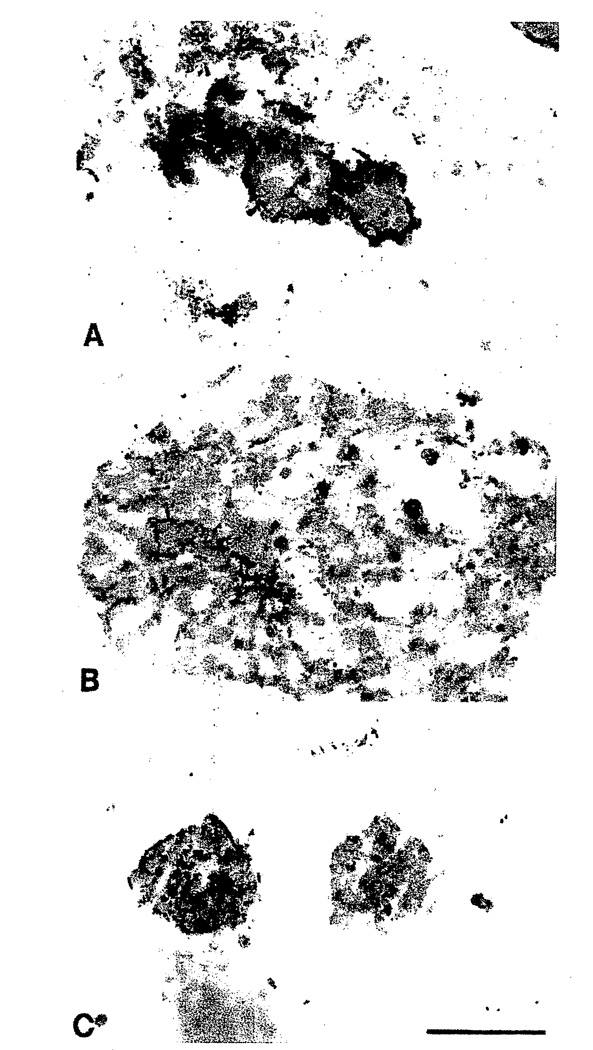
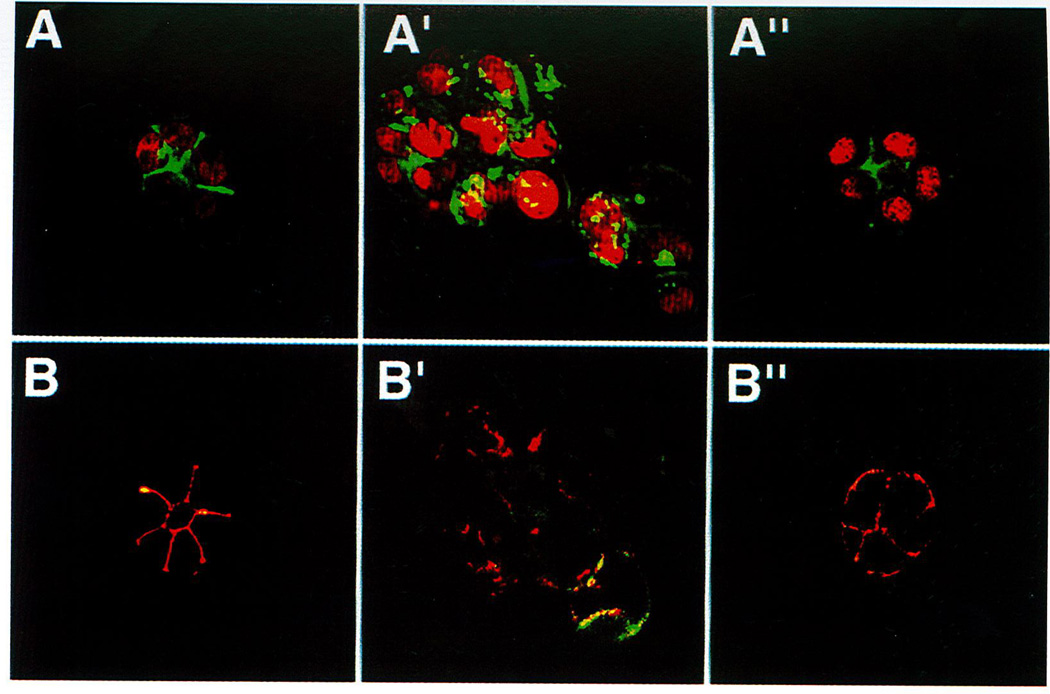
In this brief review, the development of breast cancer is discussed from the vantage of phenotypic differentiation, similar to what has been considered over the years for leukemias and melanomas, both of which express easily visible differentiation markers (Hart and Easty, 1991; Clarke et al., 1995; Lynch, 1995; Sachs, 1996; Sledge, 1996). The review is divided into a theoretical background for human breast differentiation and a discussion of recent experimental results in our laboratories with differentiation of breast epithelial cells. In the theoretical background, in situ markers of differentiation of normal breast and carcinomas are discussed with emphasis on their possible implications for tumor therapy. So far, most of the emphasis regarding differentiation therapy of tumors has been focused on the possible action of soluble factors, such as colony-stimulating factors in leukemias and retinoic acids in solid tumors (Lotan, 1996; Sachs, 1996). However, an emerging and promising new avenue in this area appears to point to additional factors, such as the cellular form and extracellular matrix (ECM) (Bissel et al., 1982; Bissel and Barcellos-Hoff, 1987; Ingber, 1992). The recent interest in these parameters has evolved along with an increasing understanding of the molecular composition of the ECM, and of the molecular basis of the classical findings that normal cell--in contrast to tumor cells--are anchorage dependent for survival and growth (Folkman and Moscona, 1978; Hannigan et al., 1996). We now know that this is the case for epithelial as well as fibroblastic cells, and that interaction with ECM is crucial for such regulation. Indeed, ECM and integrins are emerging as the central regulators of differentiation, apoptosis, and cancer (Boudreau et al., 1995; Boudreau and Bissel, 1996; Werb et al., 1996; Bissell, 1997; Weaver, et al., 1997). In the experimental part, we elaborate on our own recent experiments with functional culture models of the human breast, with particular emphasis on how "normal" and cancer cells could be defined within a reconstituted ECM. Special attention is given to integrins, the prominent ECM receptors. We further discuss a number of recent experimental results, all of which point to the same conclusion: namely that phenotypic reversion toward a more normal state for epithelial tumors is no longer an elusive goal. Thus "therapy by differentiation" could be broadened to include not only blood-borne tumors, but also solid tumors of epithelial origin.
Figures