

Tumor promoter arsenite activates extracellular signal-regulated kinase through a signaling pathway mediated by epidermal growth factor receptor and Shc
- PMID: 9710602
- PMCID: PMC109103
- DOI: 10.1128/MCB.18.9.5178
Tumor promoter arsenite activates extracellular signal-regulated kinase through a signaling pathway mediated by epidermal growth factor receptor and Shc
Abstract
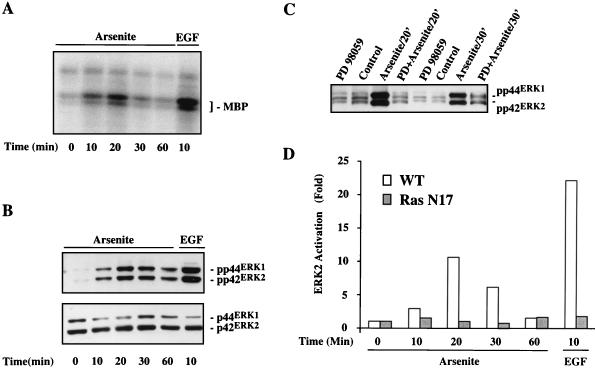
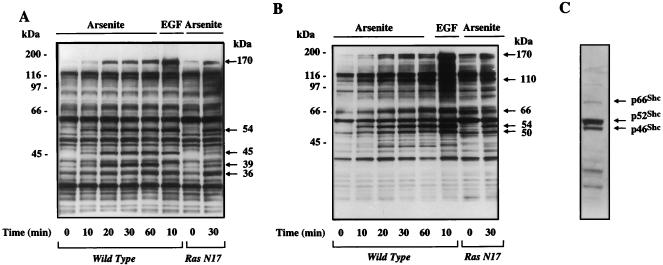
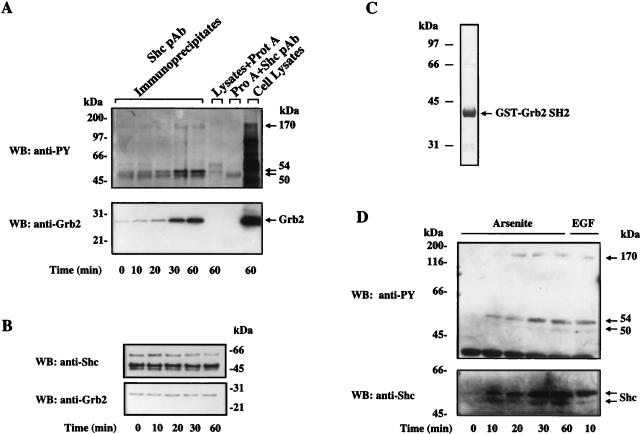
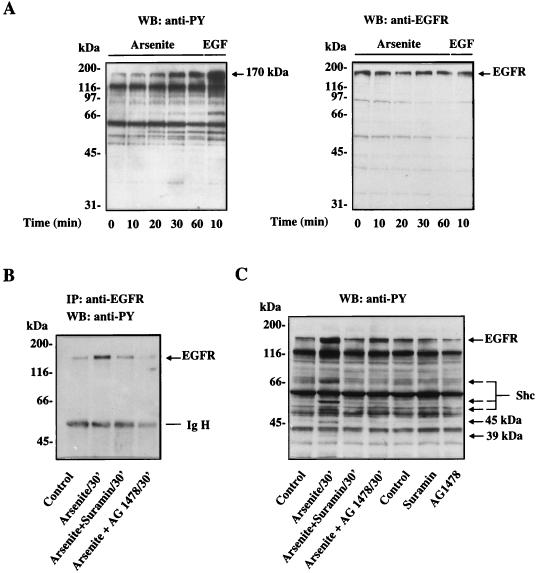
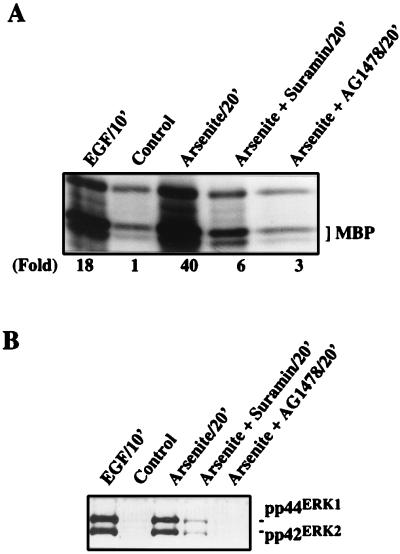
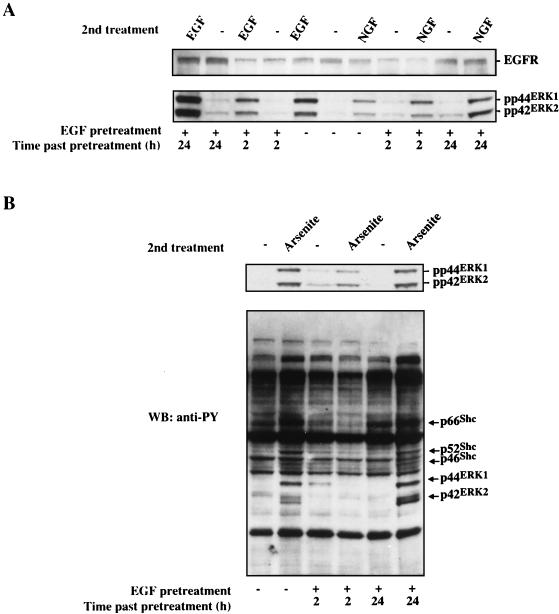
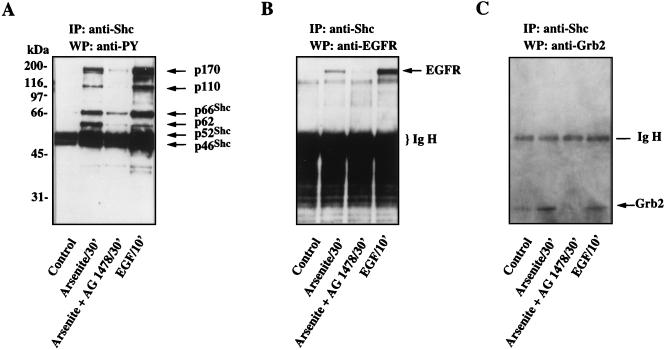
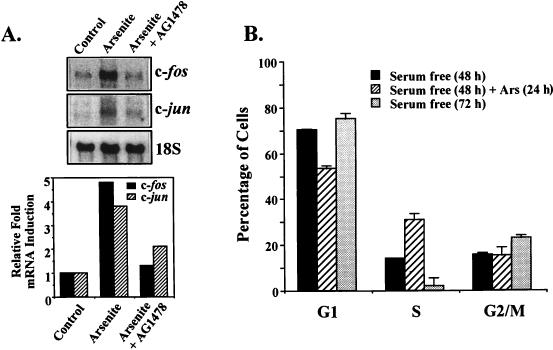
Although arsenite is an established carcinogen, the mechanisms underlying its tumor-promoting properties are poorly understood. Previously, we reported that arsenite treatment leads to the activation of the extracellular signal-regulated kinase (ERK) in rat PC12 cells through a Ras-dependent pathway. To identify potential mediators of the upstream signaling cascade, we examined the tyrosine phosphorylation profile in cells exposed to arsenite. Arsenite treatment rapidly stimulated tyrosine phosphorylation of several proteins in a Ras-independent manner, with a pattern similar to that seen in response to epidermal growth factor (EGF) treatment. Among these phosphorylated proteins were three isoforms of the proto-oncoprotein Shc as well as the EGF receptor (EGFR). Tyrosine phosphorylation of Shc allowed for enhanced interactions between Shc and Grb2 as identified by coimmunoprecipitation experiments. The arsenite-induced tyrosine phosphorylation of Shc, enhancement of Shc and Grb2 interactions, and activation of ERK were all drastically reduced by treatment of cells with either the general growth factor receptor poison suramin or the EGFR-selective inhibitor tyrphostin AG1478. Down-regulation of EGFR expression through pretreatment of cells with EGF also attenuated ERK activation and Shc tyrosine phosphorylation in response to arsenite treatment. These results demonstrate that the EGFR and Shc are critical mediators in the activation of the Ras/ERK signaling cascade by arsenite and suggest that arsenite acts as a tumor promoter largely by usurping this growth factor signaling pathway.
Figures
References
-
- Angel P, Karin M. The role of Jun, Fos and the AP-1 complex in cell-proliferation and transformation. Biochim Biophys Acta. 1991;1072:129–157. - PubMed
-
- Avruch J, Zhang X F, Kyriakis J M. Raf meets Ras: completing the framework of a signal transduction pathway. Trends Biochem Sci. 1994;19:279–283. - PubMed
-
- Bagla P, Kaiser J. India’s spreading health crisis draws global arsenic experts. Science. 1996;274:174–175. - PubMed
-
- Bonfini L, Migliaccio E, Pelicci G, Lanfrancone L, Pelicci P G. Not all Shc’s roads lead to Ras. Trends Biochem Sci. 1996;21:257–261. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous