

Dual cyclin-binding domains are required for p107 to function as a kinase inhibitor
- PMID: 9710622
- PMCID: PMC109123
- DOI: 10.1128/MCB.18.9.5380
Dual cyclin-binding domains are required for p107 to function as a kinase inhibitor
Abstract
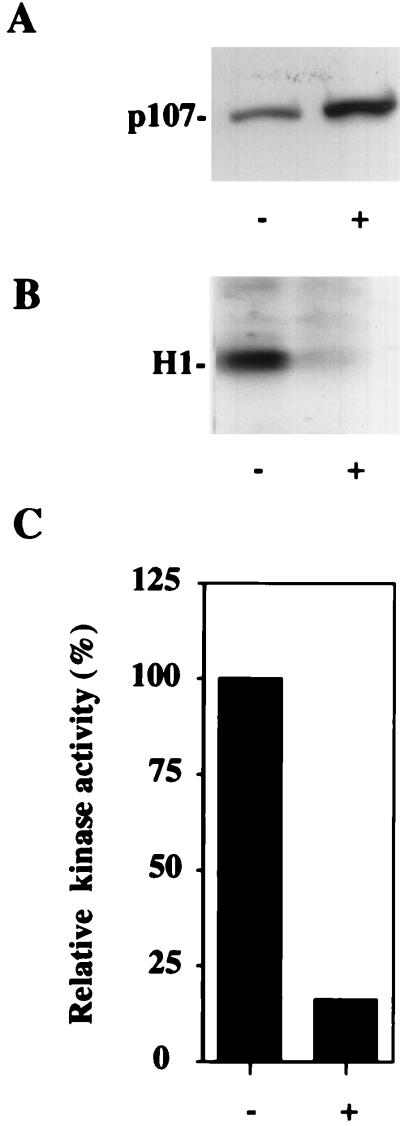
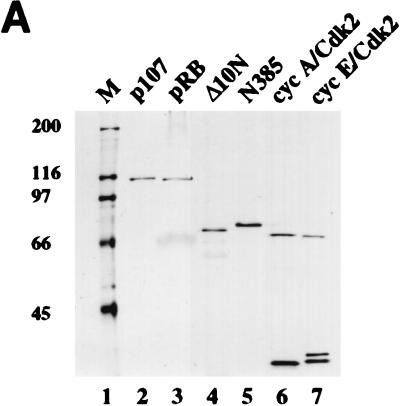
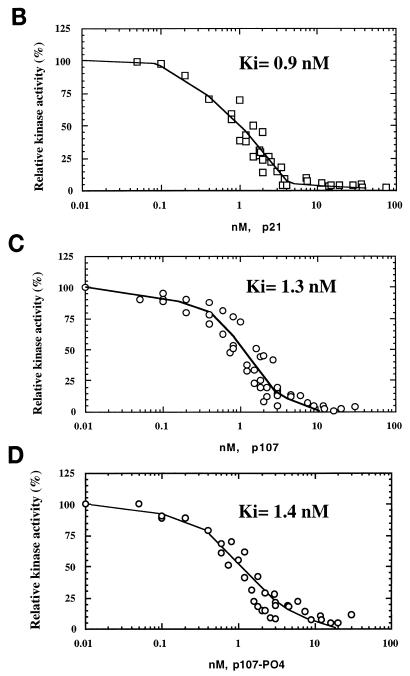
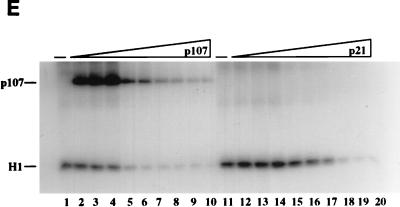
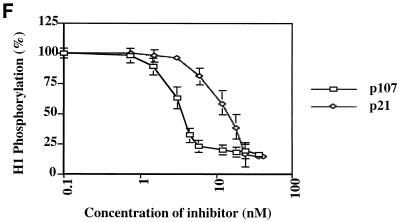
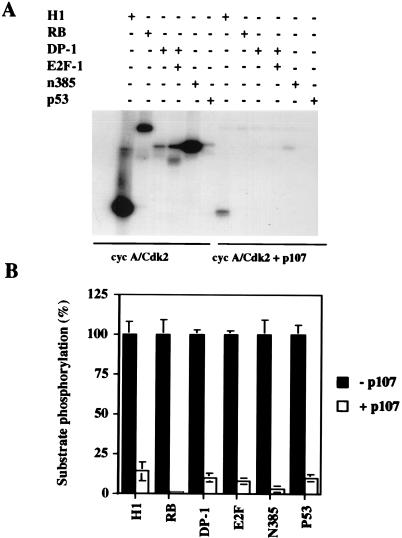
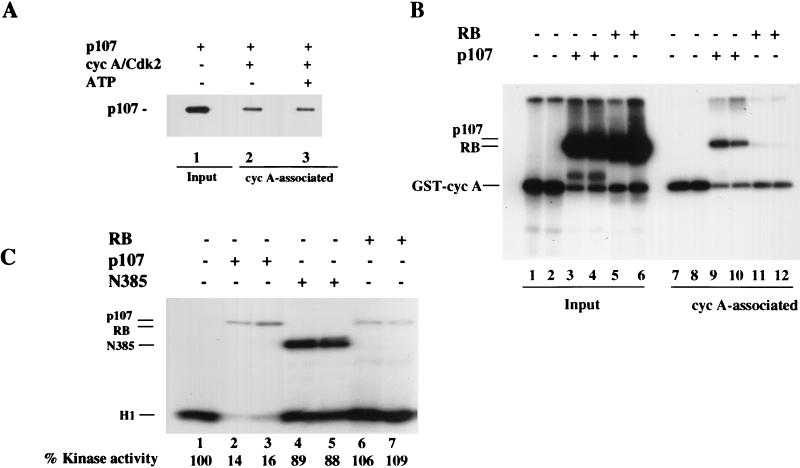
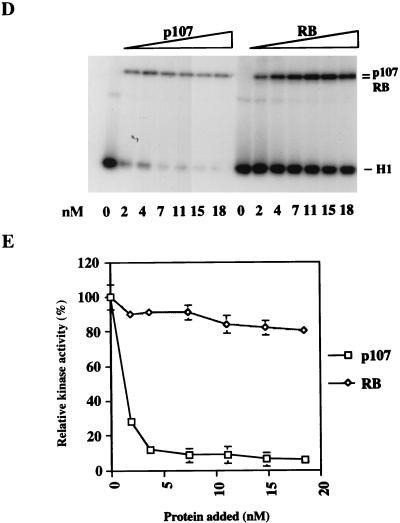
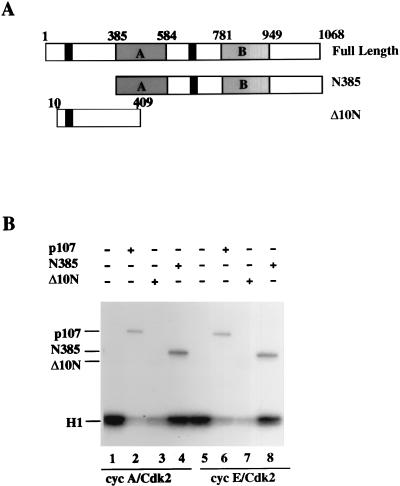
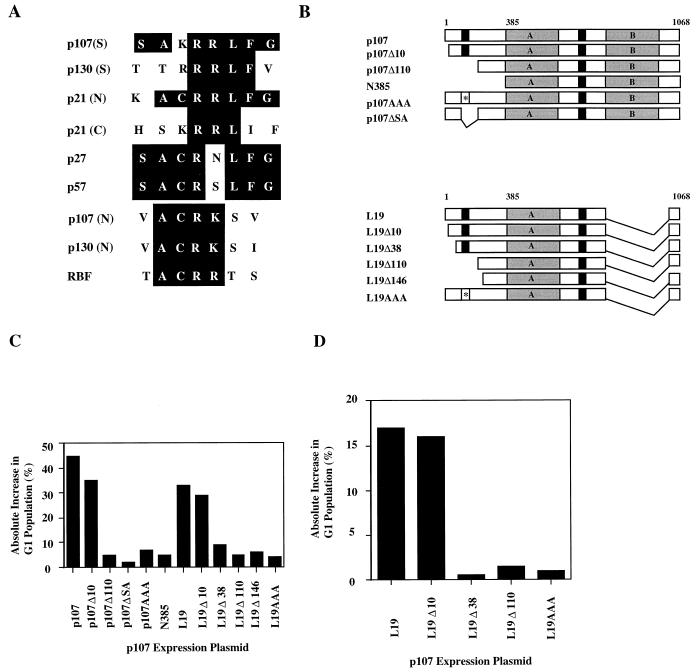
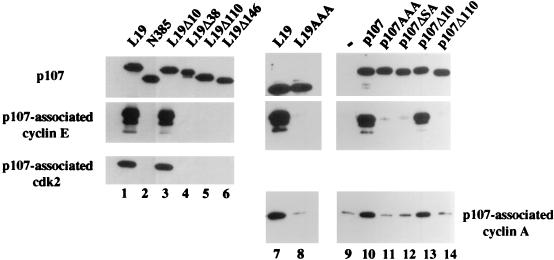
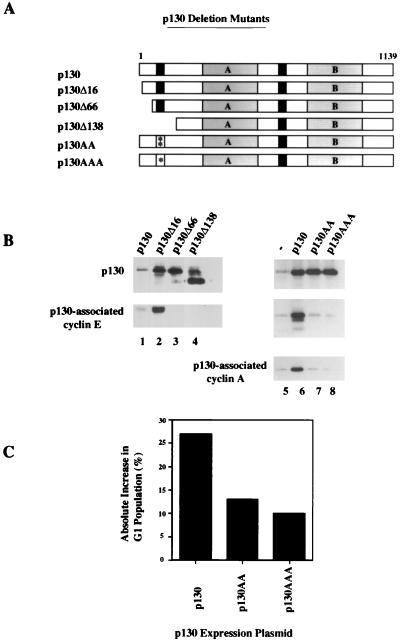
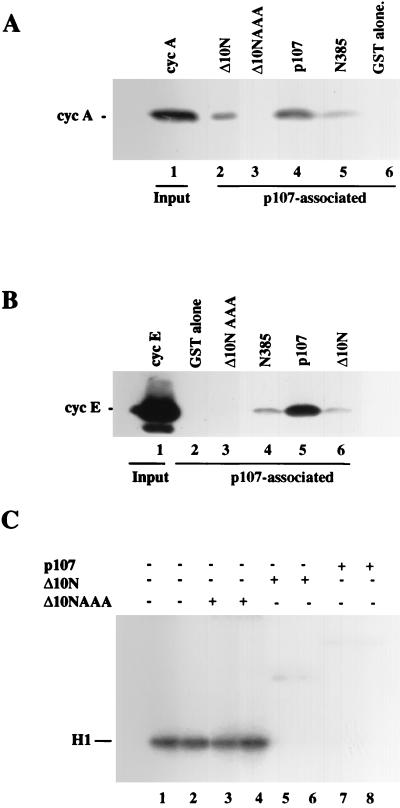
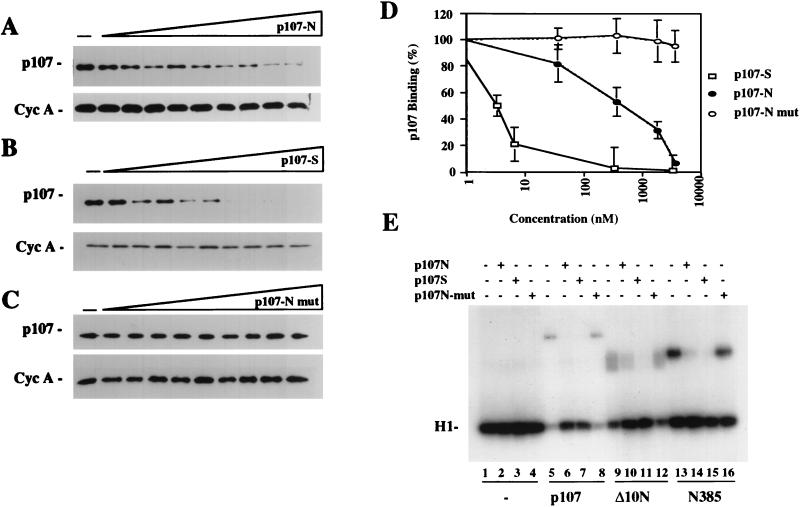
The retinoblastoma (pRB) family of proteins includes three proteins known to suppress growth of mammalian cells. Previously we had found that growth suppression by two of these proteins, p107 and p130, could result from the inhibition of associated cyclin-dependent kinases (cdks). One important unresolved issue, however, is the mechanism through which inhibition occurs. Here we present in vivo and in vitro evidence to suggest that p107 is a bona fide inhibitor of both cyclin A-cdk2 and cyclin E-cdk2 that exhibits an inhibitory constant (Ki) comparable to that of the cdk inhibitor p21/WAF1. In contrast, pRB is unable to inhibit cdks. Further reminiscent of p21, a second cyclin-binding site was mapped to the amino-terminal portions of p107 and p130. This amino-terminal domain is capable of inhibiting cyclin-cdk2 complexes, although it is not a potent substrate for these kinases. In contrast, a carboxy-terminal fragment of p107 that contains the previously identified cyclin-binding domain serves as an excellent kinase substrate although it is unable to inhibit either kinase. Clustered point mutations suggest that the amino-terminal domain is functionally important for cyclin binding and growth suppression. Moreover, peptides spanning the cyclin-binding region are capable of interfering with p107 binding to cyclin-cdk2 complexes and kinase inhibition. Our ability to distinguish between p107 and p130 as inhibitors rather than simple substrates suggests that these proteins may represent true inhibitors of cdks.
Figures
References
-
- Beijersbergen R L, Carlee L, Verkhoven R M, Bernards R. Regulation of the retinoblastoma protein-related p107 by G1 cyclin complexes. Genes Dev. 1995;9:1340–1353. - PubMed
-
- Cao L, Faha B, Dembski M, Tsai L-H, Harlow E, Dyson N. Independent binding of the retinoblastoma protein and p107 to the transcription factor E2F. Nature. 1992;355:176–179. - PubMed
-
- Chen I-T, Akamatsu M, Smith M L, Lung F-D T, Duba D, Roller P P, Fornace A J, O’Connor P M. Characterization of p21cip1/waf1 peptide domains required for cyclin E/cdk2 and PCNA interactions. Oncogene. 1996;12:595–607. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources