

Acute Plasmodium chabaudi chabaudi malaria infection induces antibodies which bind to the surfaces of parasitized erythrocytes and promote their phagocytosis by macrophages in vitro
- PMID: 9712751
- PMCID: PMC108489
- DOI: 10.1128/IAI.66.9.4080-4086.1998
Acute Plasmodium chabaudi chabaudi malaria infection induces antibodies which bind to the surfaces of parasitized erythrocytes and promote their phagocytosis by macrophages in vitro
Abstract
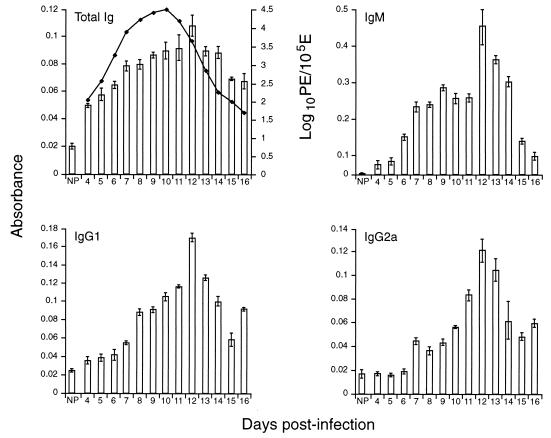
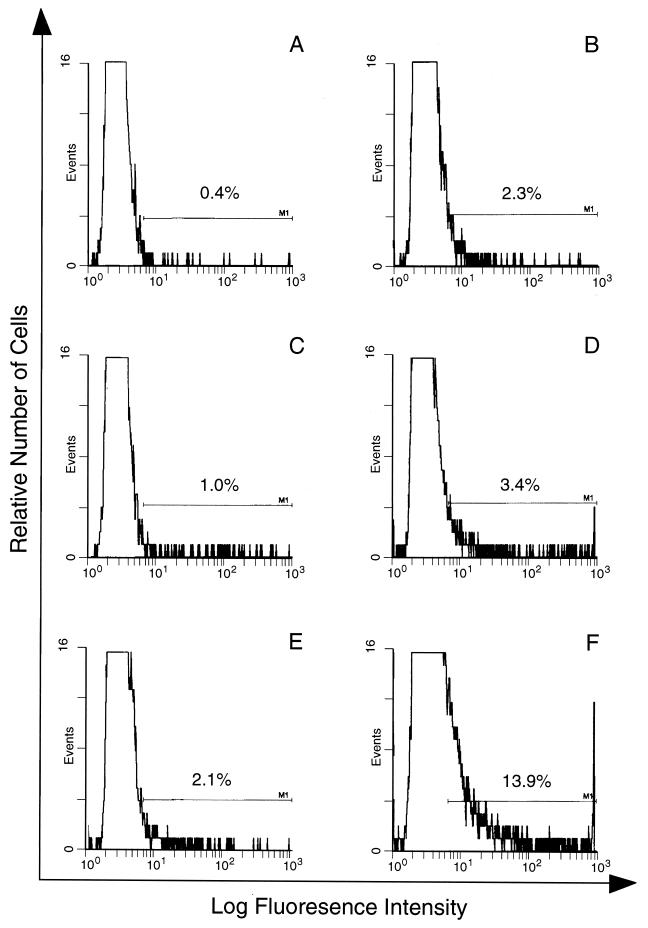
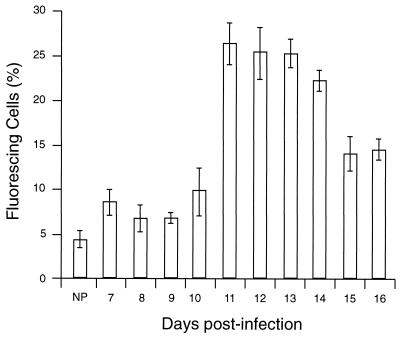
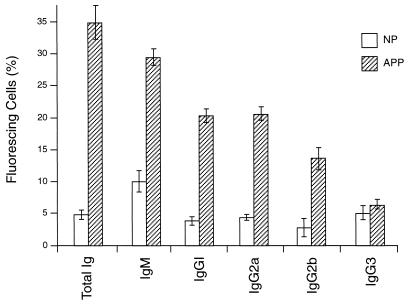
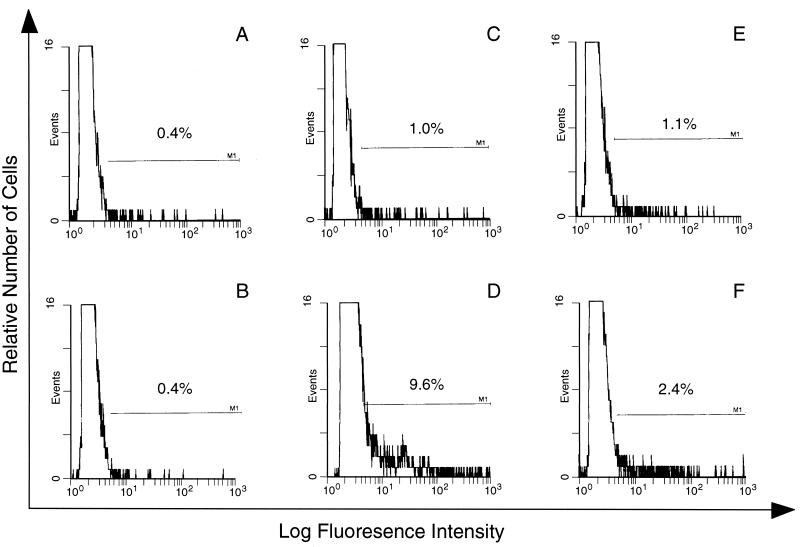
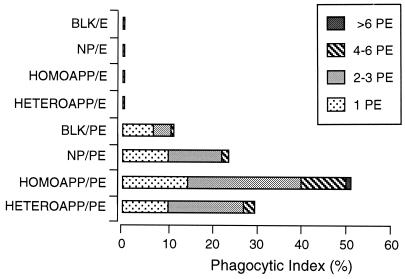
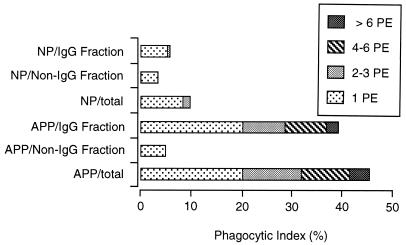
CBA/Ca mice infected with 5 x 10(4) Plasmodium chabaudi chabaudi AS-parasitized erythrocytes experience acute but self-limiting infections of relatively short duration. Parasitemia peaks ( approximately 40% infected erythrocytes) on day 10 or 11 and is then partially resolved over the ensuing 5 to 6 days, a period referred to as crisis. How humoral and cellular immune mechanisms contribute to parasite killing and/or clearance during crisis is controversial. Humoral immunity might be parasite variant, line, or species specific, while cellular immune responses would be relatively less specific. For P. c. chabaudi AS, parasite clearance is largely species and line specific during this time, which suggests a primary role for antibody activity. Accordingly, acute-phase plasma (APP; taken from P. c. chabaudi AS-infected mice at day 11 or 12 postinfection) was examined for the presence of parasite-specific antibody activity by enzyme-linked immunosorbent assay. Antibody binding to the surface of intact, live parasitized erythrocytes, particularly those containing mature (trophozoite and schizont) parasites, was demonstrated by immunofluorescence in APP and the immunoglobulin G (IgG)-containing fraction thereof. Unfractionated APP (from P. c. chabaudi AS-infected mice), as well as its IgG fraction, specifically mediated the opsonization and internalization of P. c. chabaudi AS-parasitized erythrocytes by macrophages in vitro. APP from another parasite line (P. c. chabaudi CB) did not mediate the same effect against P. c. chabaudi AS-parasitized erythrocytes. These results, which may represent one mechanism of parasite removal during crisis, are discussed in relation to the parasite variant, line, and species specificity of parasite clearance during this time.
Figures
References
-
- Baird J, Purnomo K, Basri H, Bangs M J, Andersen E M, Jones T R, Masbar S, Harjosuwarno S, Subianto B, Arbani P R. Age-specific prevalence of Plasmodium falciparum among six populations with limited histories of exposure to endemic malaria. Am J Trop Med Hyg. 1993;49:707–719. - PubMed
-
- Baruch D I, Pasloske B L, Singh H B, Bi X, Ma X C, Feldman M, Taraschi T F, Howard R J. Cloning the P. falciparum gene encoding PfEMP1, a malarial variant antigen and adherence receptor on the surface of parasitized human erythrocytes. Cell. 1995;82:77–87. - PubMed
-
- Brannan L R, Turner C M, Phillips R S. Malaria parasites undergo antigenic variation at high rates in vivo. Proc R Soc Lond Ser B. 1994;256:71–75. - PubMed
-
- Greenwood B M, Bradley A K, Greenwood A M, Byass P, Jammeh K, Marsh K, Tulloch S, Oldfield F S, Hayes R. Mortality and morbidity from malaria among children in a rural area of The Gambia, West Africa. Trans R Soc Trop Med Hyg. 1987;81:478–486. - PubMed
-
- Howard R J, Barnwell J W, Rock E P, Neequaye J, Ofori-Adjei D, Maloy W L, Lyon J A, Saul A. Two approximately 300 kilodalton Plasmodium falciparum proteins at the surface membrane of infected erythrocytes. Mol Biochem Parasitol. 1988;27:207–223. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
