

Identification of the gene encoding the alternative sigma factor sigmaB from Listeria monocytogenes and its role in osmotolerance
- PMID: 9721294
- PMCID: PMC107466
- DOI: 10.1128/JB.180.17.4547-4554.1998
Identification of the gene encoding the alternative sigma factor sigmaB from Listeria monocytogenes and its role in osmotolerance
Abstract
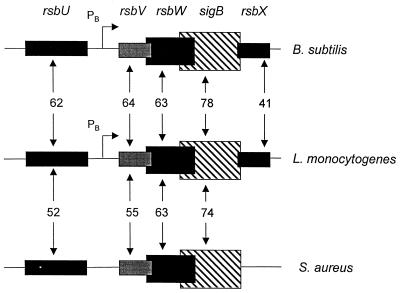
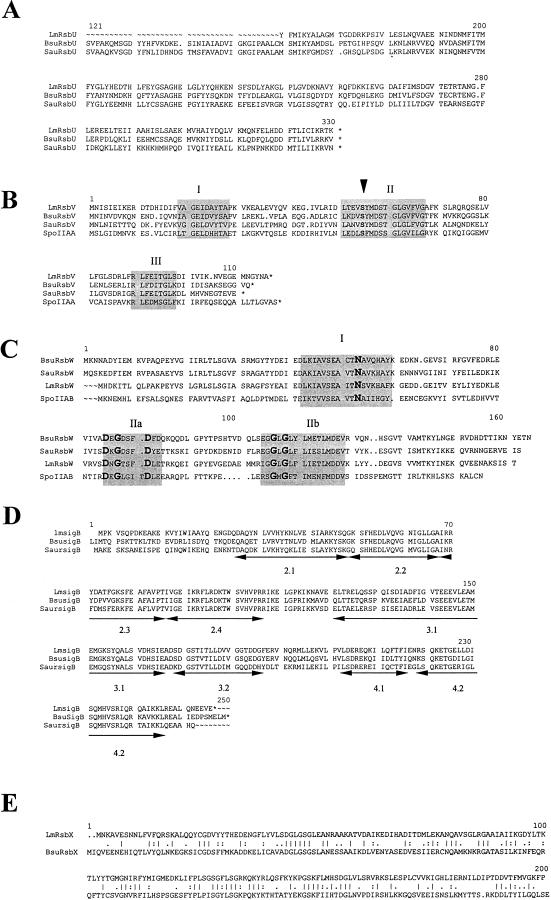
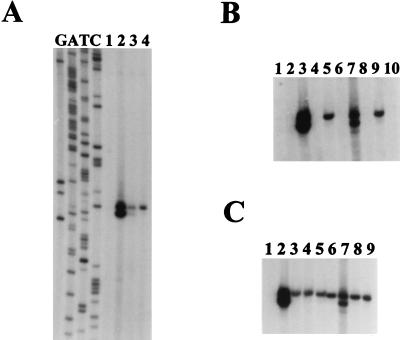
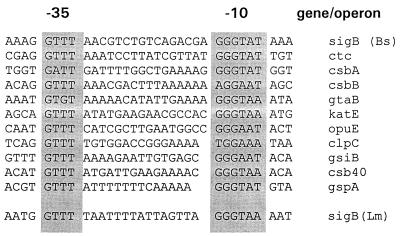
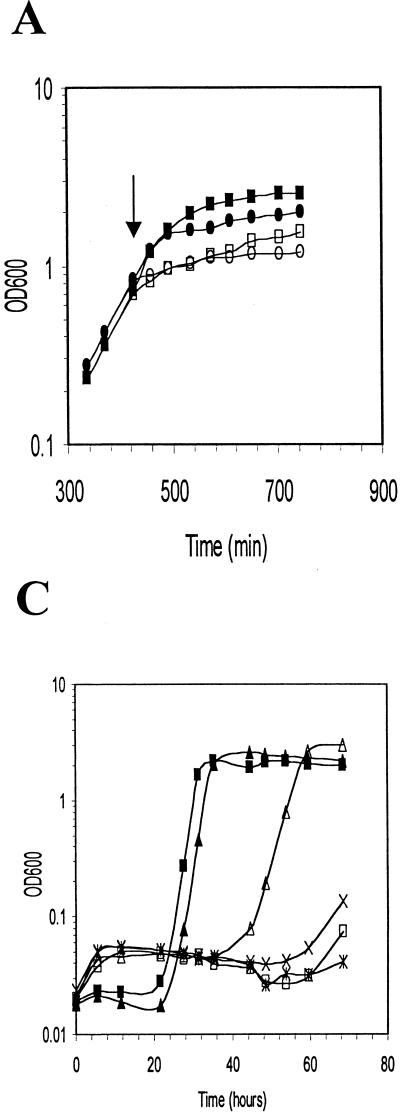
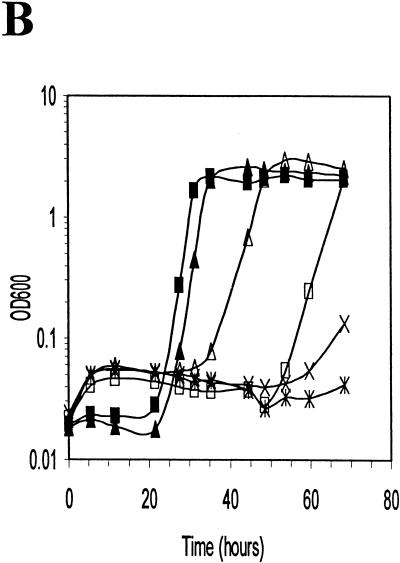
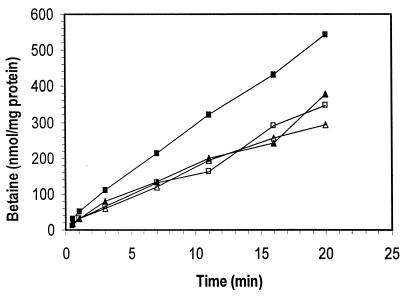
Listeria monocytogenes is well known for its robust physiology, which permits growth at low temperatures under conditions of high osmolarity and low pH. Although studies have provided insight into the mechanisms used by L. monocytogenes to allay the physiological consequences of these adverse environments, little is known about how these responses are coordinated. In the studies presented here, we have cloned the sigB gene and several rsb genes from L. monocytogenes, encoding homologs of the alternative sigma factor sigmaB and the RsbUVWX proteins, which govern transcription of a general stress regulon in the related bacterium Bacillus subtilis. The L. monocytogenes and B. subtilis sigB and rsb genes are similar in sequence and physical organization; however, we observed that the activity of sigmaB in L. monocytogenes was uniquely responsive to osmotic upshifting, temperature downshifting, and the presence of EDTA in the growth medium. The magnitude of the response was greatest after an osmotic upshift, suggesting a role for sigmaB in coordinating osmotic responses in L. monocytogenes. A null mutation in the sigB gene led to substantial defects in the ability of L. monocytogenes to use betaine and carnitine as osmoprotectants. Subsequent measurements of betaine transport confirmed that the absence of sigmaB reduced the ability of the cells to accumulate betaine. Thus, sigmaB coordinates responses to a variety of physical and chemical signals, and its function facilitates the growth of L. monocytogenes under conditions of high osmotic strength.
Figures
References
-
- Akbar S, Kang C M, Gaidenko T A, Price C W. Modulator protein RsbR regulates environmental signalling in the general stress pathway of Bacillus subtilis. Mol Microbiol. 1997;24:567–578. - PubMed
-
- Alper S, Duncan L, Losick R. An adenosine nucleotide switch controlling the activity of a cell type-specific transcription factor in Bacillus subtilis. Cell. 1994;77:195–205. - PubMed
-
- Becker, L. A., and A. K. Benson. 1998. Unpublished data.
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
