

Molecular basis for the dominant white phenotype in the domestic pig
- PMID: 9724328
- PMCID: PMC310759
- DOI: 10.1101/gr.8.8.826
Molecular basis for the dominant white phenotype in the domestic pig
Abstract
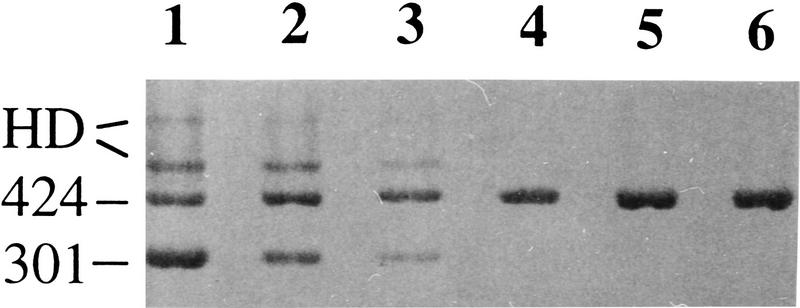
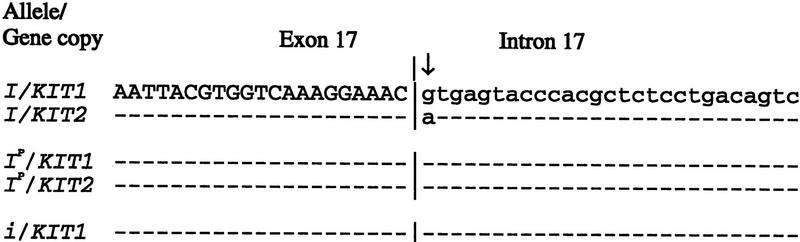
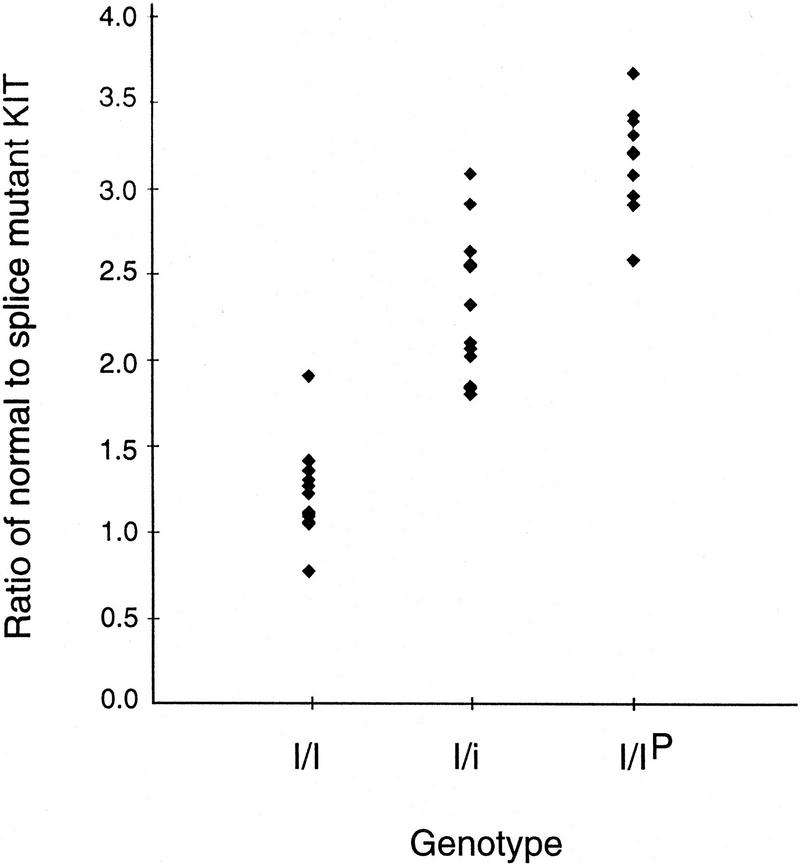
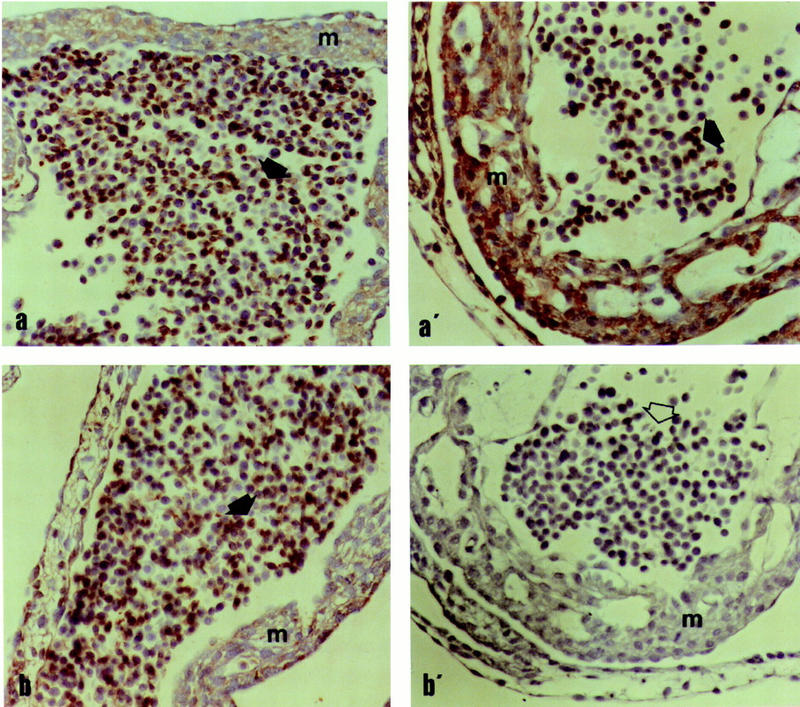
The change of phenotypic traits in domestic animals and crops as a response to selective breeding mimics the much slower evolutionary change in natural populations. Here, we describe that the dominant white phenotype in domestic pigs is caused by two mutations in the KIT gene encoding the mast/stem cell growth factor receptor (MGF), one gene duplication associated with a partially dominant phenotype and a splice mutation in one of the copies leading to the fully dominant allele. The splice mutation is a G to A substitution in the first nucleotide of intron 17 and leads to skipping of exon 17. The duplication is most likely a regulatory mutation affecting KIT expression, whereas the splice mutation is expected to cause a receptor with impaired or absent tyrosine kinase activity. Immunocytochemistry showed that this variant form is expressed in 17- to 19-day-old pig embryos. Hundreds of millions of white pigs around the world are assumed to be heterozygous or homozygous for the two mutations. [The EMBL accession numbers for porcine KIT1*0101, KIT1*0202, KIT2*0202, and KIT2*0101 are AJ223228-AJ223231, respectively.]
Figures
References
-
- Anonymous. SAS v. 6.10. Cary, NC: SAS Institute Inc.; 1990.
-
- Chabot B, Stephensson DA, Chapman VM, Besmer P, Bernstein A. The proto-oncogene c-kit encoding a transmembrane tyrosine kinase receptor maps to the mouse W locus. Nature. 1988;335:88–89. - PubMed
-
- Deol MS. The relationship between abnormalities of pigmentation and of the inner ear. Proc Roy Soc (London) A. 1970;175:201–217. - PubMed
-
- Duttlinger R, Manova K, Berrozpe G, Chu TY, DeLeon V, Timokhina I, Chaganti RS, Zelenetz AD, Bachvarova RF, Besmer P. The Wsh and Ph mutations affect the c-kit expression profile: c-kit misexpression in embryogenesis impairs melanogenesis in Wsh and Ph mutant mice. Proc Natl Acad Sci. 1995;92:3754–3758. - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources