

Protein kinase A-catalyzed phosphorylation of heat shock protein 60 chaperone regulates its attachment to histone 2B in the T lymphocyte plasma membrane
- PMID: 9724719
- PMCID: PMC27910
- DOI: 10.1073/pnas.95.18.10425
Protein kinase A-catalyzed phosphorylation of heat shock protein 60 chaperone regulates its attachment to histone 2B in the T lymphocyte plasma membrane
Abstract
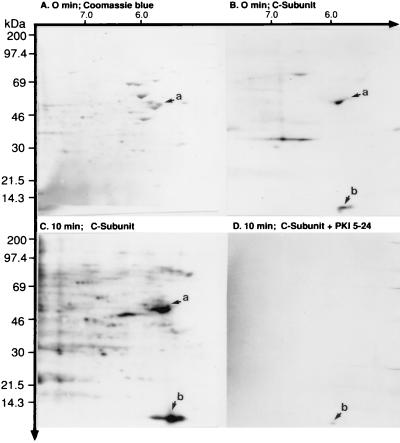
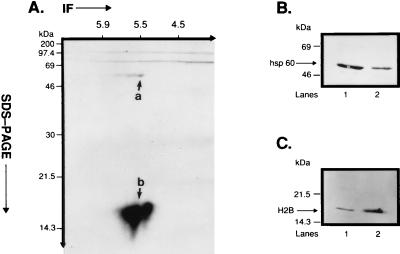
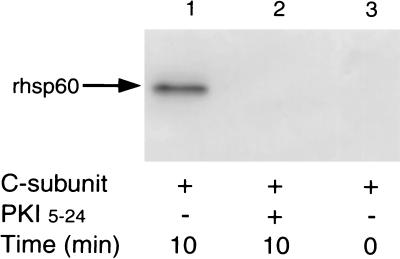
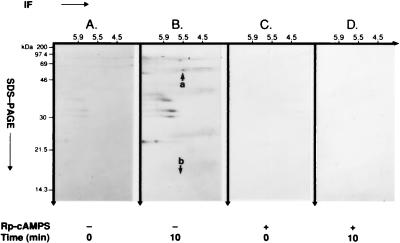
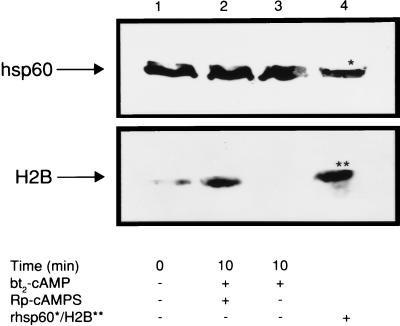
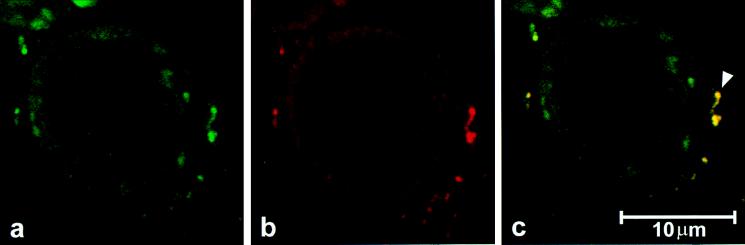
Accumulating evidence suggests that the mitochondrial molecular chaperone heat shock protein 60 (hsp60) also can localize in extramitochondrial sites. However, direct evidence that hsp60 functions as a chaperone outside of mitochondria is presently lacking. A 60-kDa protein that is present in the plasma membrane of a human leukemic CD4(+) CEM-SS T cell line and is phosphorylated by protein kinase A (PKA) was identified as hsp60. An 18-kDa plasma membrane-associated protein coimmunoprecipitated with hsp60 and was identified as histone 2B (H2B). Hsp60 physically associated with H2B when both molecules were in their dephospho forms. By contrast, PKA-catalyzed phosphorylation of both hsp60 and H2B caused dissociation of H2B from hsp60 and loss of H2B from the plasma membrane of intact T cells. These results suggest that (i) hsp60 and H2B can localize in the T cell plasma membrane; (ii) hsp60 functions as a molecular chaperone for H2B; and (iii) PKA-catalyzed phosphorylation of both hsp60 and H2B appears to regulate the attachment of H2B to hsp60. We propose a model in which phosphorylation/dephosphorylation regulates chaperoning of H2B by hsp60 in the plasma membrane.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
