

Multiple independent origins of mitochondrial gene order in birds
- PMID: 9724766
- PMCID: PMC27957
- DOI: 10.1073/pnas.95.18.10693
Multiple independent origins of mitochondrial gene order in birds
Abstract
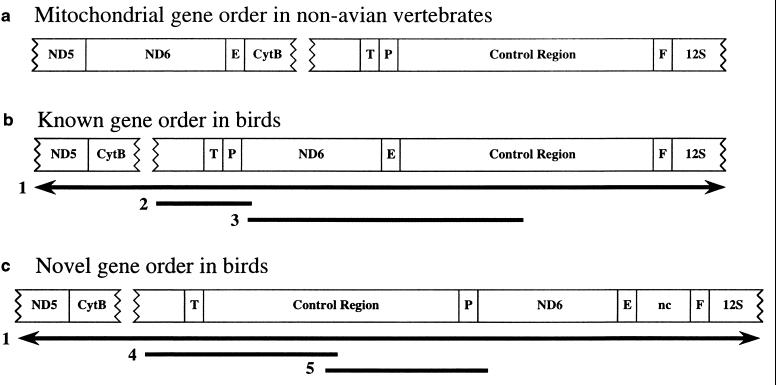
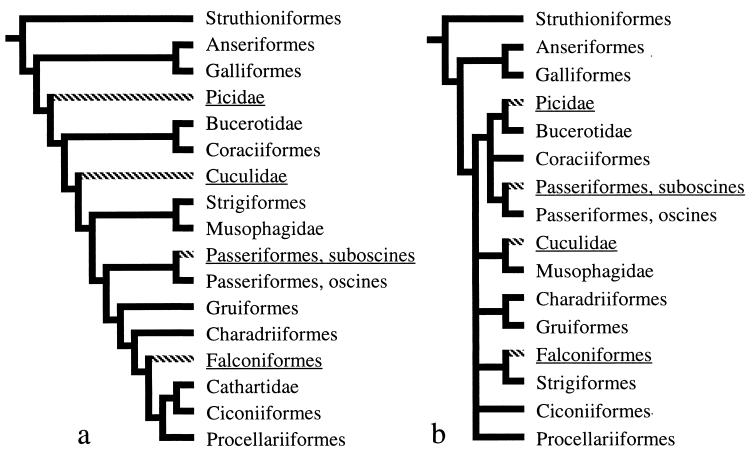
Mitochondrial genomes of all vertebrate animals analyzed to date have the same 37 genes, whose arrangement in the circular DNA molecule varies only in the relative position of a few genes. This relative conservation suggests that mitochondrial gene order characters have potential utility as phylogenetic markers for higher-level vertebrate taxa. We report discovery of a mitochondrial gene order that has had multiple independent originations within birds, based on sampling of 137 species representing 13 traditionally recognized orders. This provides evidence of parallel evolution in mitochondrial gene order for animals. Our results indicate operation of physical constraints on mitochondrial gene order changes and support models for gene order change based on replication error. Bird mitochondria have a displaced OL (origin of light-strand replication site) as do various other Reptilia taxa prone to gene order changes. Our findings point to the need for broad taxonomic sampling in using mitochondrial gene order for phylogenetic analyses. We found, however, that the alternative mitochondrial gene orders distinguish the two primary groups of songbirds (order Passeriformes), oscines and suboscines, in agreement with other molecular as well as morphological data sets. Thus, although mitochondrial gene order characters appear susceptible to some parallel evolution because of mechanistic constraints, they do hold promise for phylogenetic studies.
Figures
References
-
- Boore J L, Collins T M, Stanton D, Daehler L L, Brown W M. Nature (London) 1995;376:163–165. - PubMed
-
- Quinn T W, Mindell D P. Mol Phylogenet Evol. 1996;5:344–351. - PubMed
-
- Macey J R, Larson A, Ananjeva N B, Fang Z, Papenfuss T J. Mol Biol Evol. 1997;14:91–104. - PubMed
-
- Pääbo S, Thomas W K, Whitfield K M, Kumazawa Y, Wilson A C. J Mol Evol. 1991;33:426–430. - PubMed
LinkOut - more resources
Full Text Sources
