

Model of sarcomeric Ca2+ movements, including ATP Ca2+ binding and diffusion, during activation of frog skeletal muscle
- PMID: 9725890
- PMCID: PMC2229419
- DOI: 10.1085/jgp.112.3.297
Model of sarcomeric Ca2+ movements, including ATP Ca2+ binding and diffusion, during activation of frog skeletal muscle
Abstract
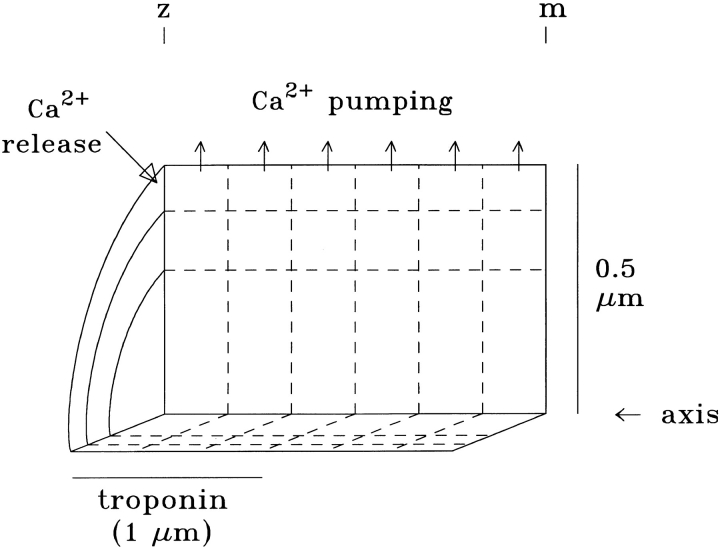
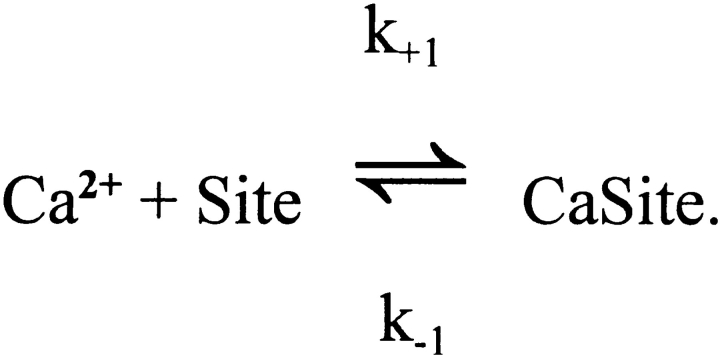
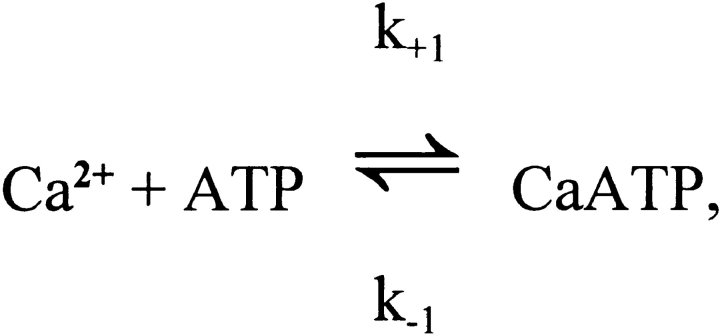
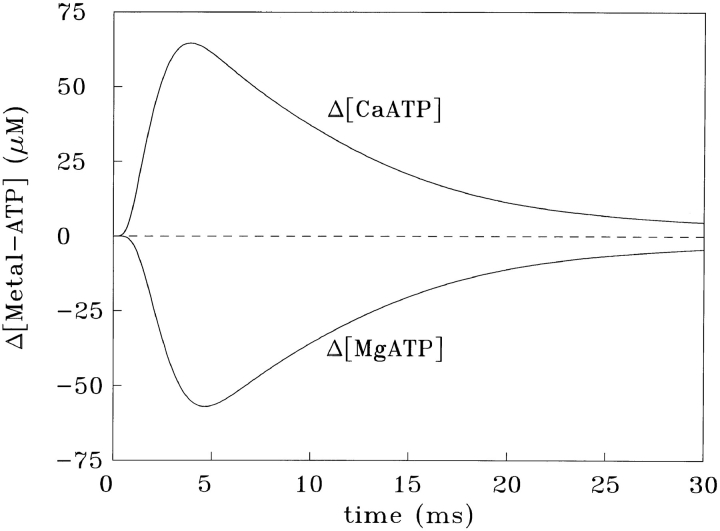
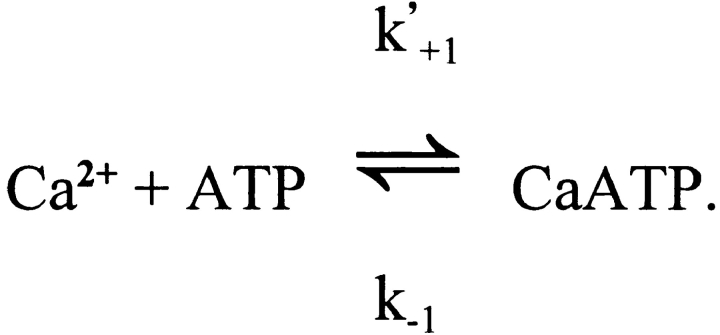
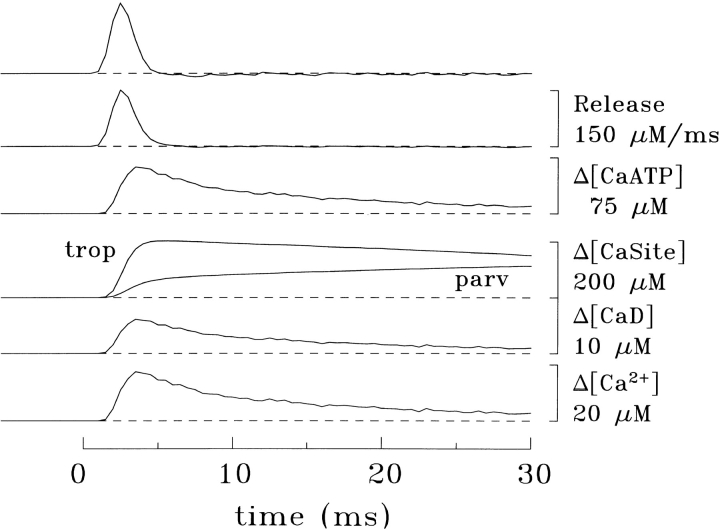
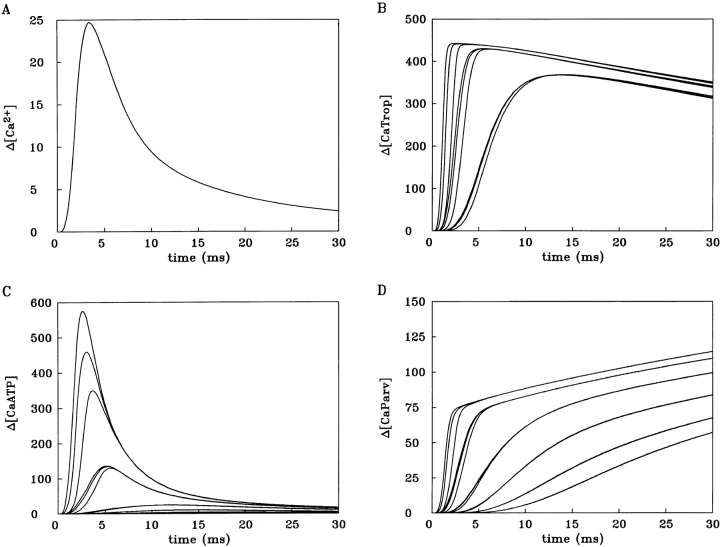
Cannell and Allen (1984. Biophys. J. 45:913-925) introduced the use of a multi-compartment model to estimate the time course of spread of calcium ions (Ca2+) within a half sarcomere of a frog skeletal muscle fiber activated by an action potential. Under the assumption that the sites of sarcoplasmic reticulum (SR) Ca2+ release are located radially around each myofibril at the Z line, their model calculated the spread of released Ca2+ both along and into the half sarcomere. During diffusion, Ca2+ was assumed to react with metal-binding sites on parvalbumin (a diffusible Ca2+- and Mg2+-binding protein) as well as with fixed sites on troponin. We have developed a similar model, but with several modifications that reflect current knowledge of the myoplasmic environment and SR Ca2+ release. We use a myoplasmic diffusion constant for free Ca2+ that is twofold smaller and an SR Ca2+ release function in response to an action potential that is threefold briefer than used previously. Additionally, our model includes the effects of Ca2+ and Mg2+ binding by adenosine 5'-triphosphate (ATP) and the diffusion of Ca2+-bound ATP (CaATP). Under the assumption that the total myoplasmic concentration of ATP is 8 mM and that the amplitude of SR Ca2+ release is sufficient to drive the peak change in free [Ca2+] (Delta[Ca2+]) to 18 microM (the approximate spatially averaged value that is observed experimentally), our model calculates that (a) the spatially averaged peak increase in [CaATP] is 64 microM; (b) the peak saturation of troponin with Ca2+ is high along the entire thin filament; and (c) the half-width of Delta[Ca2+] is consistent with that observed experimentally. Without ATP, the calculated half-width of spatially averaged Delta[Ca2+] is abnormally brief, and troponin saturation away from the release sites is markedly reduced. We conclude that Ca2+ binding by ATP and diffusion of CaATP make important contributions to the determination of the amplitude and the time course of Delta[Ca2+].
Figures
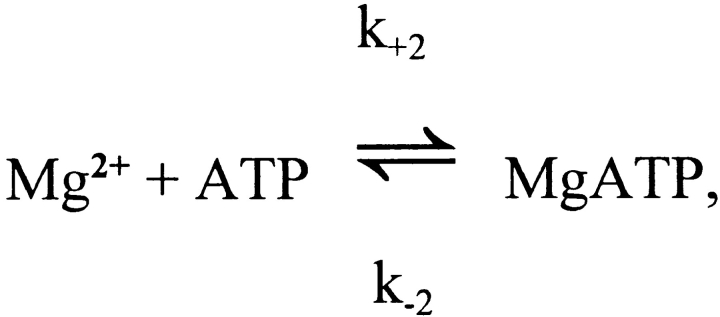


References
-
- Baylor SM, Harkins AB, Kurebayashi N. Response to Westerblad and Allen. Biophys J. 1994;66:927–928.
-
- Baylor SM, Hollingworth S. The transient binding of calcium to ATP and diffusion of Ca-ATP help shape the amplitude and time course of the myoplasmic free Ca transient (Δ[Ca]) Biophys J. 1998;74:A235.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
