

A novel synaptobrevin/VAMP homologous protein (VAMP5) is increased during in vitro myogenesis and present in the plasma membrane
- PMID: 9725904
- PMCID: PMC25509
- DOI: 10.1091/mbc.9.9.2423
A novel synaptobrevin/VAMP homologous protein (VAMP5) is increased during in vitro myogenesis and present in the plasma membrane
Abstract
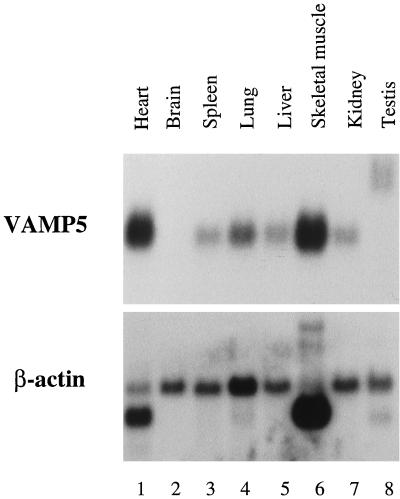
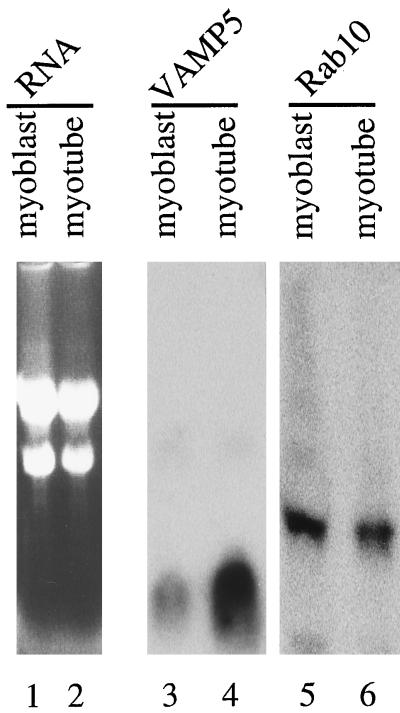
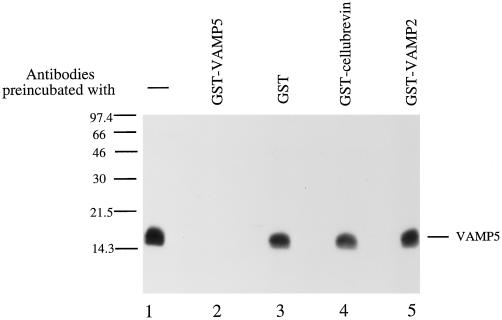
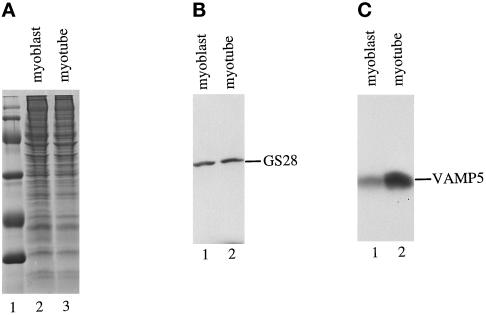
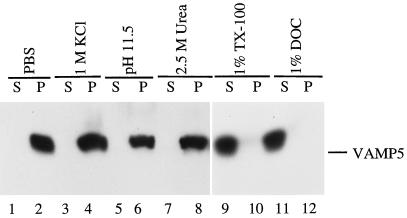
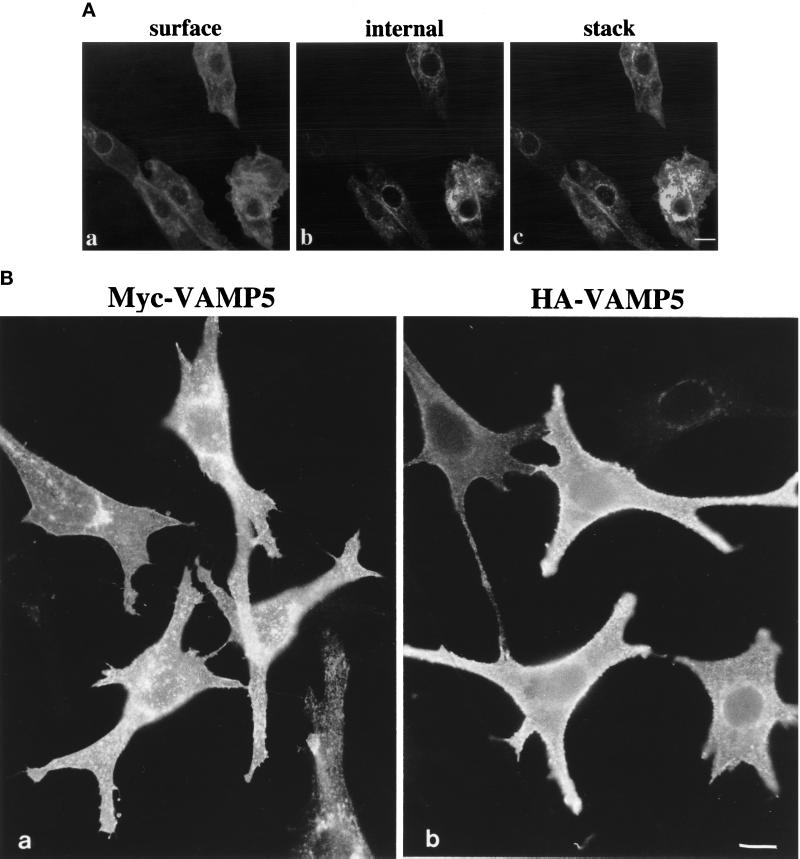
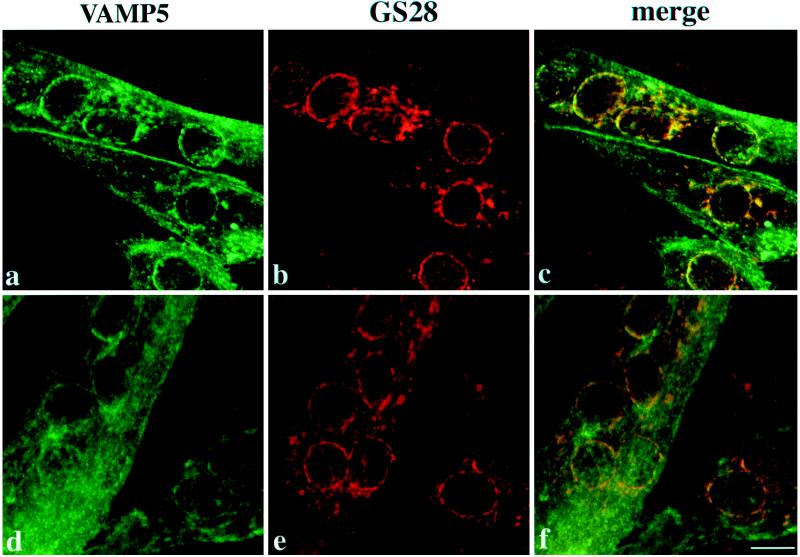
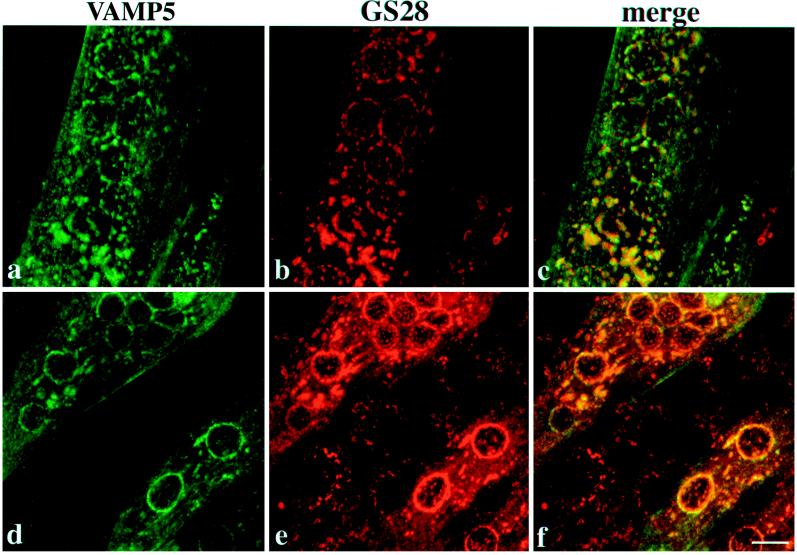
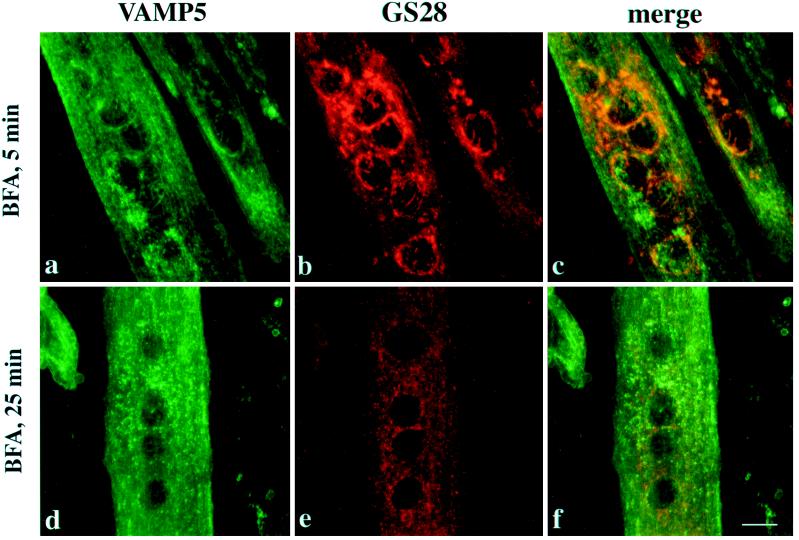
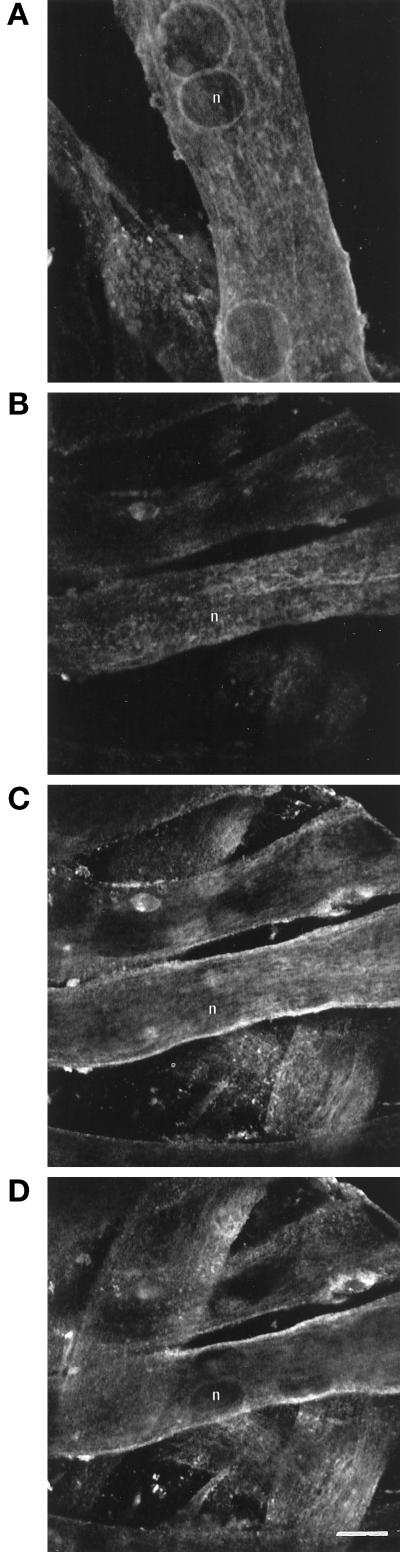
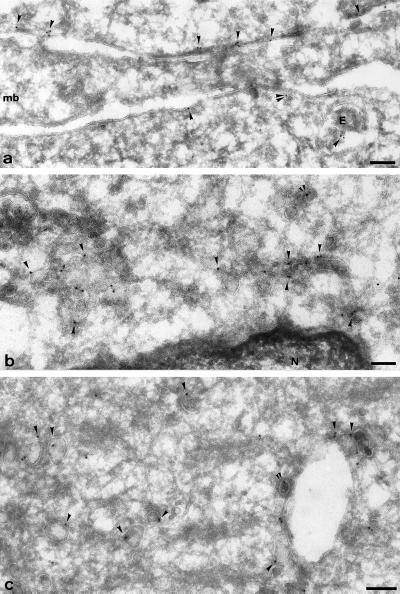
cDNA clones encoding a novel protein (VAMP5) homologous to synaptobrevins/VAMPs are detected during database searches. The predicted 102-amino acid VAMP5 harbors a 23-residue hydrophobic region near the carboxyl terminus and exhibits an overall amino acid identity of 33% with synaptobrevin/VAMP1 and 2 and cellubrevin. Northern blot analysis reveals that the mRNA for VAMP5 is preferentially expressed in the skeletal muscle and heart, whereas significantly lower levels are detected in several other tissues but not in the brain. During in vitro differentiation (myogenesis) of C2C12 myoblasts into myotubes, the mRNA level for VAMP5 is increased approximately 8- to 10-fold. Immunoblot analysis using antibodies specific for VAMP5 shows that the protein levels are also elevated approximately 6-fold during in vitro myogenesis of C2C12 cells. Indirect immunofluorescence microscopy and immunoelectron microscopy reveal that VAMP5 is associated with the plasma membrane as well as intracellular perinuclear and peripheral vesicular structures of myotubes. Epitope-tagged versions of VAMP5 are similarly targeted to the plasma membrane.
Figures

References
-
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol. 1990;215:403–410. - PubMed
-
- Bennett MK, Calakos N, Scheller RH. Syntaxin: a synaptic protein implicated in docking of synaptic vesicles at presynaptic active zones. Science. 1992;257:255–259. - PubMed
-
- Bennett MK, Garcia-Arraras JE, Elferink LA, Peterson K, Fleming AM, Hazuka CD, Scheller RH. The syntaxin family of vesicular transport receptors. Cell. 1993;74:863–873. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
