

Aerobic anoxygenic phototrophic bacteria
- PMID: 9729607
- PMCID: PMC98932
- DOI: 10.1128/MMBR.62.3.695-724.1998
Aerobic anoxygenic phototrophic bacteria
Abstract
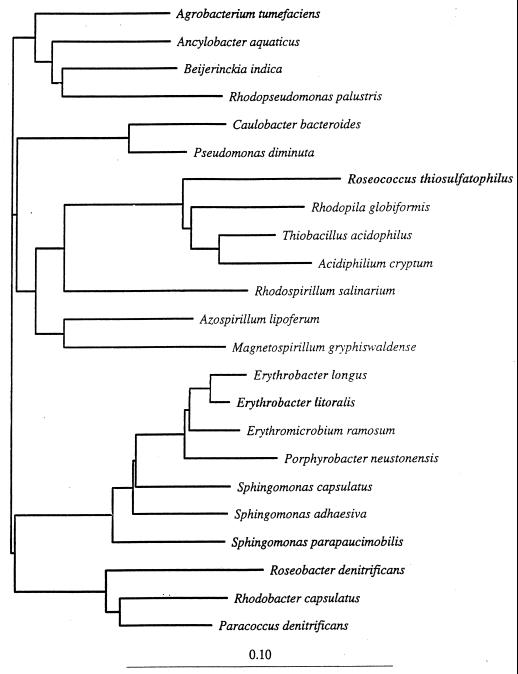
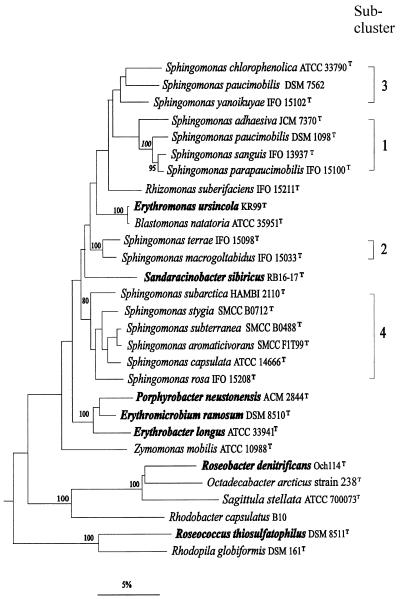
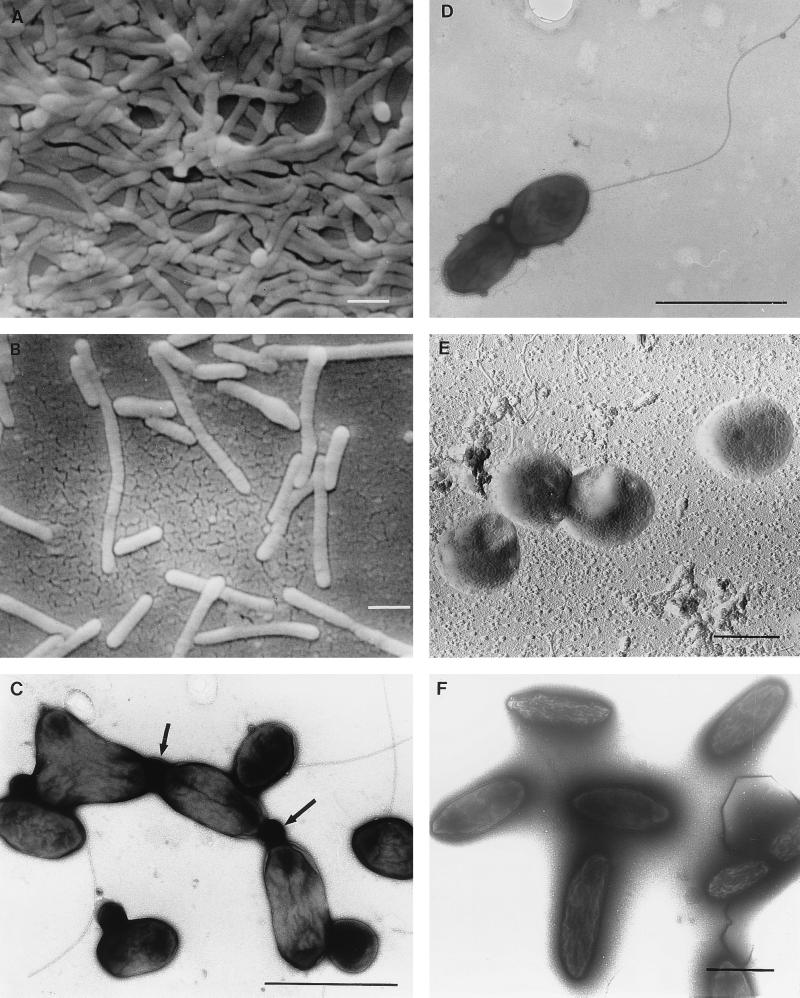
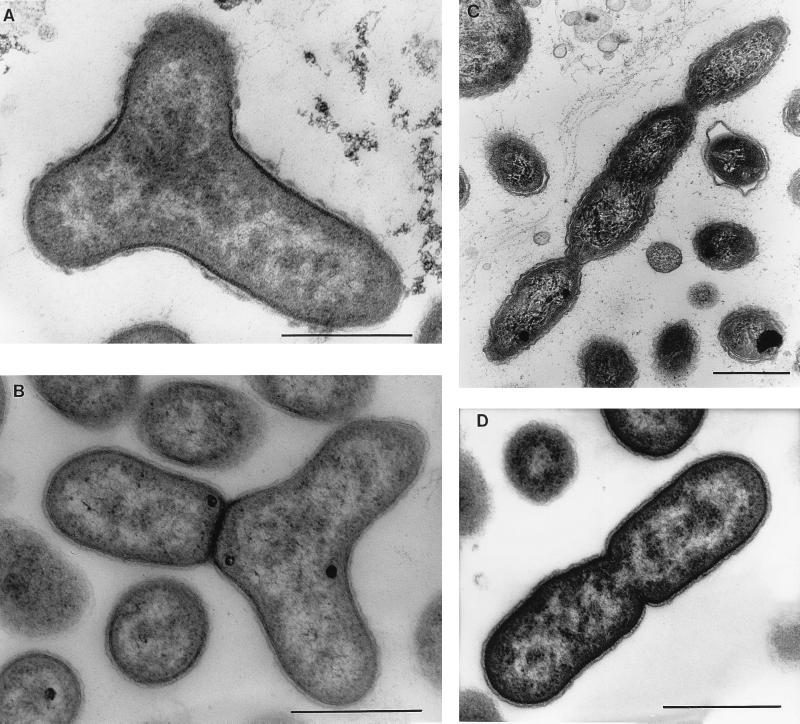
The aerobic anoxygenic phototrophic bacteria are a relatively recently discovered bacterial group. Although taxonomically and phylogenetically heterogeneous, these bacteria share the following distinguishing features: the presence of bacteriochlorophyll a incorporated into reaction center and light-harvesting complexes, low levels of the photosynthetic unit in cells, an abundance of carotenoids, a strong inhibition by light of bacteriochlorophyll synthesis, and the inability to grow photosynthetically under anaerobic conditions. Aerobic anoxygenic phototrophic bacteria are classified in two marine (Erythrobacter and Roseobacter) and six freshwater (Acidiphilium, Erythromicrobium, Erythromonas, Porphyrobacter, Roseococcus, and Sandaracinobacter) genera, which phylogenetically belong to the alpha-1, alpha-3, and alpha-4 subclasses of the class Proteobacteria. Despite this phylogenetic information, the evolution and ancestry of their photosynthetic properties are unclear. We discuss several current proposals for the evolutionary origin of aerobic phototrophic bacteria. The closest phylogenetic relatives of aerobic phototrophic bacteria include facultatively anaerobic purple nonsulfur phototrophic bacteria. Since these two bacterial groups share many properties, yet have significant differences, we compare and contrast their physiology, with an emphasis on morphology and photosynthetic and other metabolic processes.
Figures
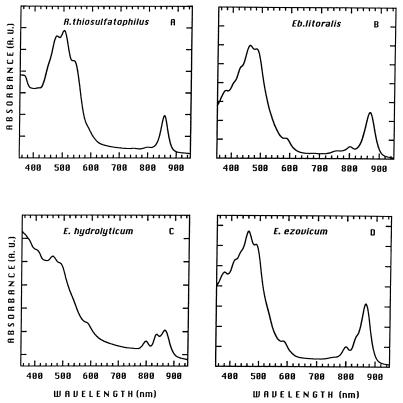

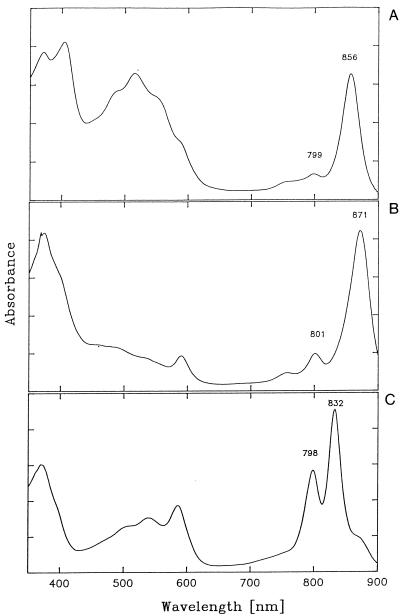
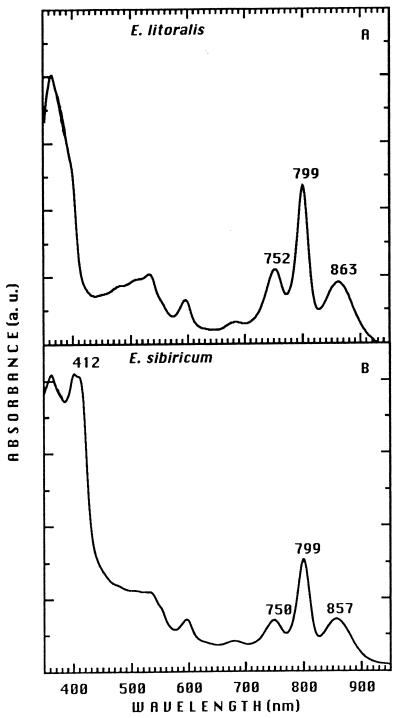
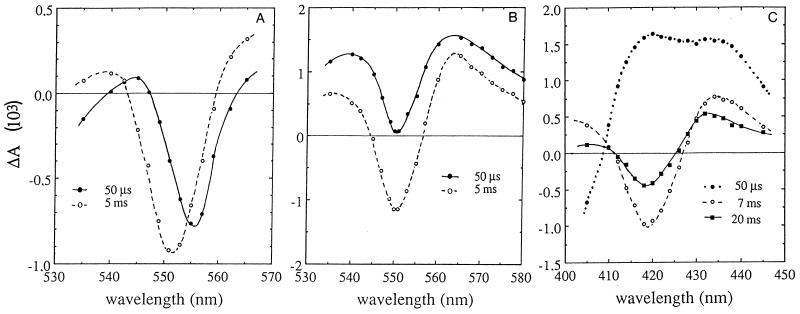
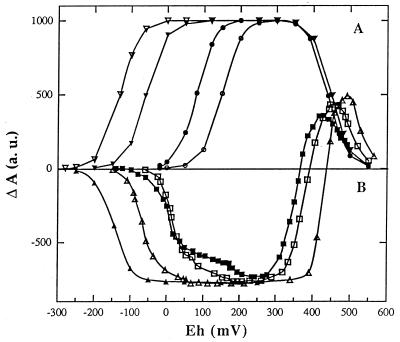
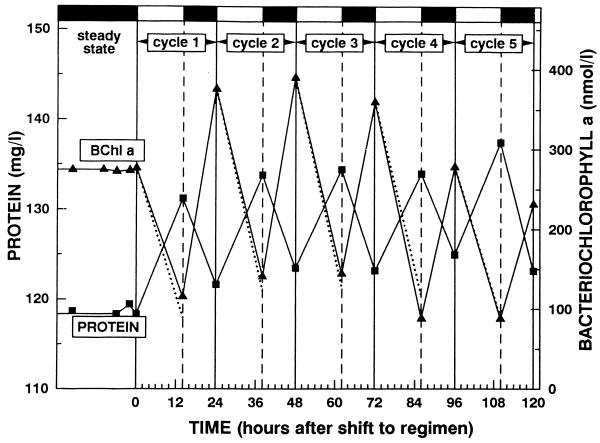
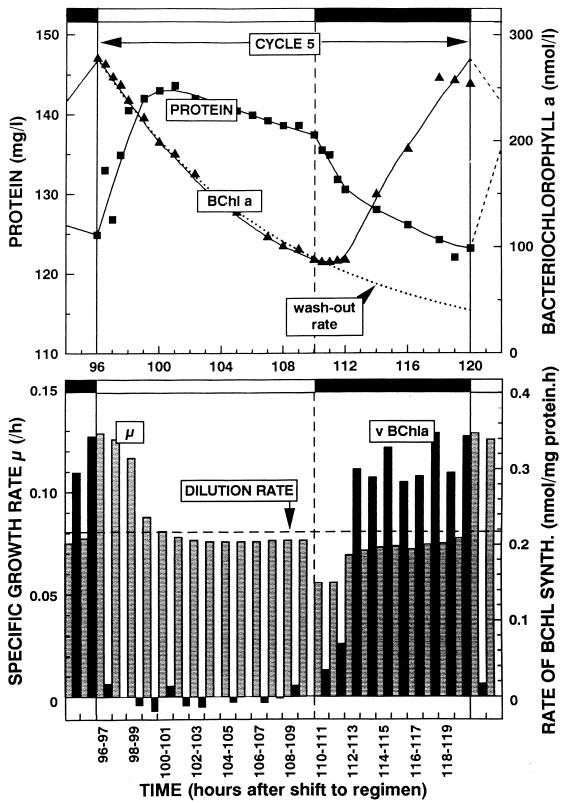
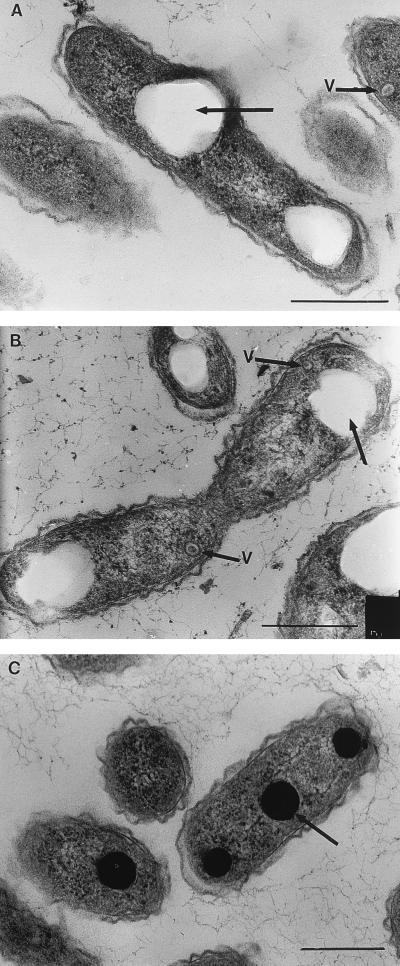

References
-
- Aagaard J, Sistrom W R. Control of synthesis of reaction center bacteriochlorophyll in photosynthetic bacteria. Photochem Photobiol. 1972;15:209–225. - PubMed
-
- Albers H, Gottschalk G. Acetate metabolism in Rhodopseudomonas gelatinosa and several other Rhodospirillaceae. Arch Microbiol. 1976;111:45–49. - PubMed
-
- Arata H, Serikawa Y, Takamiya K I. Trimethylamine-N-oxide respiration by aerobic photosynthetic bacterium, Erythrobacter sp. OCh114. J Biochem. 1988;103:1011–1015. - PubMed
-
- Armstrong G A. Genetic analysis and regulation of carotenoid biosynthesis: structure and function of the crt genes and gene products. In: Blankenship R E, Madigan M T, Bauer C E, editors. Anoxygenic photosynthetic bacteria. Dordrecht, The Netherlands: Kluwer Academic Publishers; 1995. pp. 1135–1157.
-
- Bagnall K W. Selenium, tellurium, and polonium. In: Schmidt M, Siebert W, Bagnall K W, editors. The chemistry of sulfur, selenium, tellurium and polonium. New York, N.Y: Pergamon Press; 1975. pp. 935–1008.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
