

Insertion sequences
- PMID: 9729608
- PMCID: PMC98933
- DOI: 10.1128/MMBR.62.3.725-774.1998
Insertion sequences
Abstract
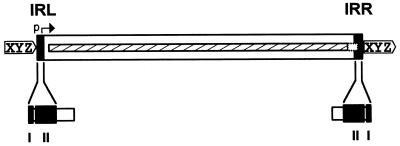
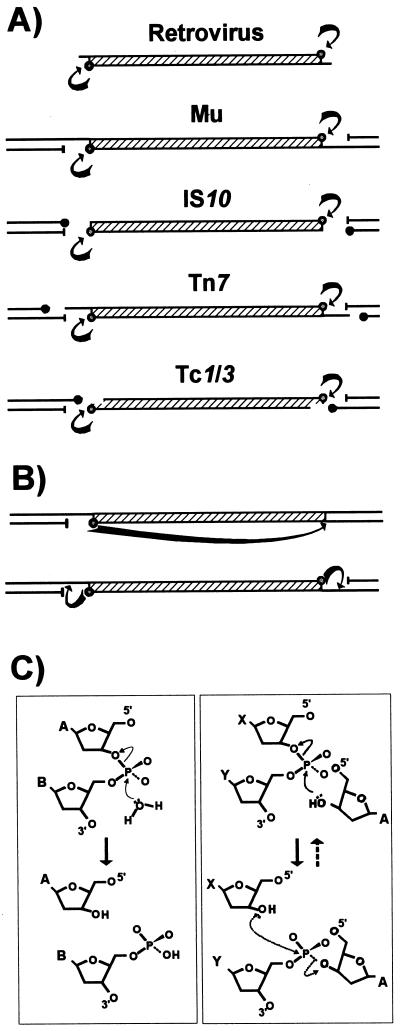
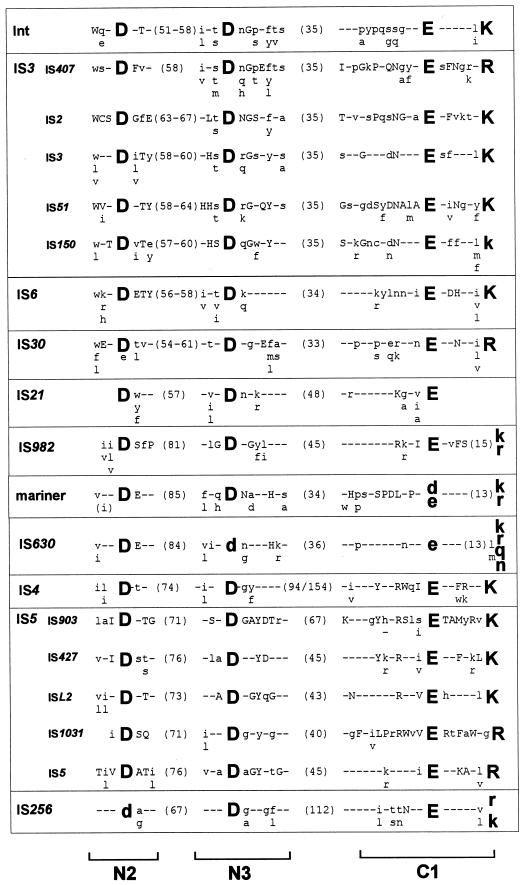
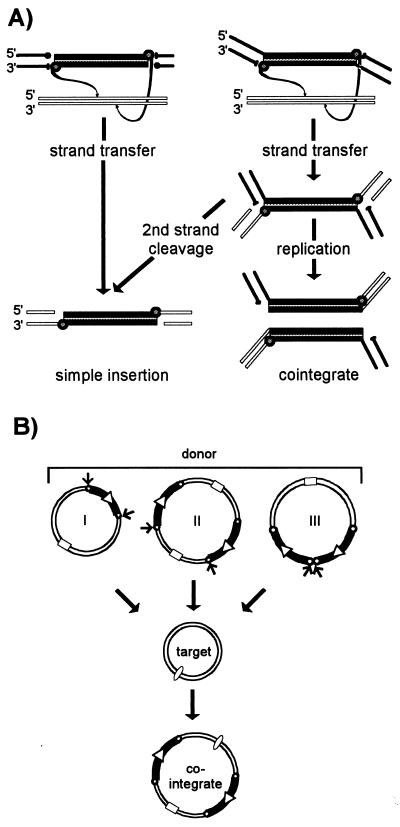
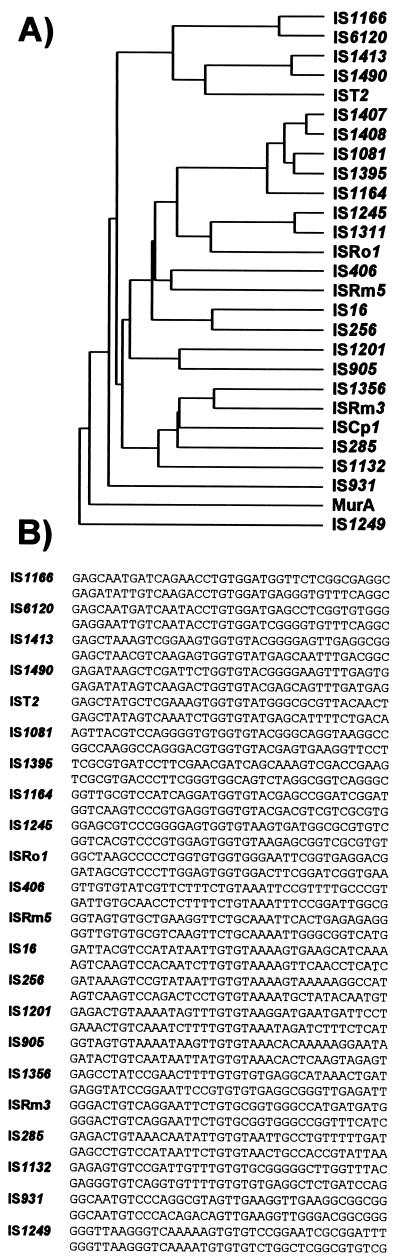
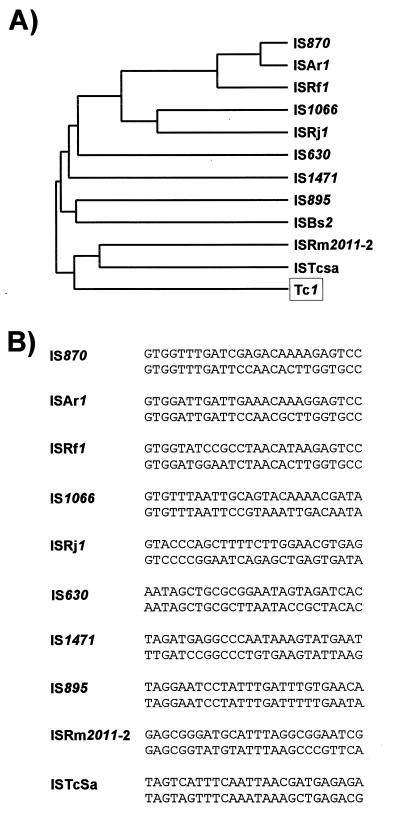
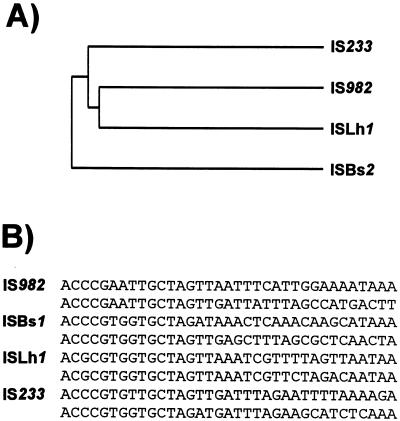
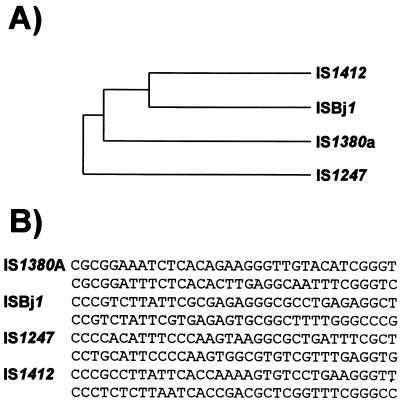
Insertion sequences (ISs) constitute an important component of most bacterial genomes. Over 500 individual ISs have been described in the literature to date, and many more are being discovered in the ongoing prokaryotic and eukaryotic genome-sequencing projects. The last 10 years have also seen some striking advances in our understanding of the transposition process itself. Not least of these has been the development of various in vitro transposition systems for both prokaryotic and eukaryotic elements and, for several of these, a detailed understanding of the transposition process at the chemical level. This review presents a general overview of the organization and function of insertion sequences of eubacterial, archaebacterial, and eukaryotic origins with particular emphasis on bacterial elements and on different aspects of the transposition mechanism. It also attempts to provide a framework for classification of these elements by assigning them to various families or groups. A total of 443 members of the collection have been grouped in 17 families based on combinations of the following criteria: (i) similarities in genetic organization (arrangement of open reading frames); (ii) marked identities or similarities in the enzymes which mediate the transposition reactions, the recombinases/transposases (Tpases); (iii) similar features of their ends (terminal IRs); and (iv) fate of the nucleotide sequence of their target sites (generation of a direct target duplication of determined length). A brief description of the mechanism(s) involved in the mobility of individual ISs in each family and of the structure-function relationships of the individual Tpases is included where available.
Figures
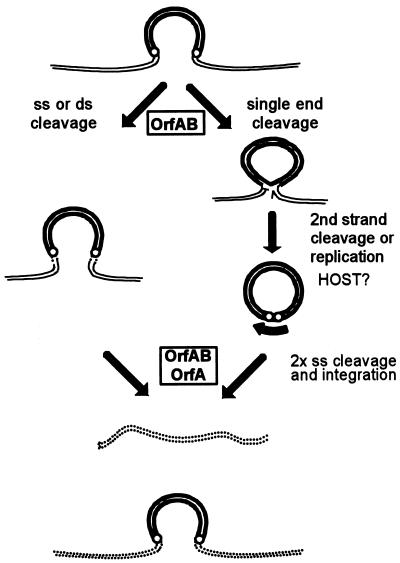

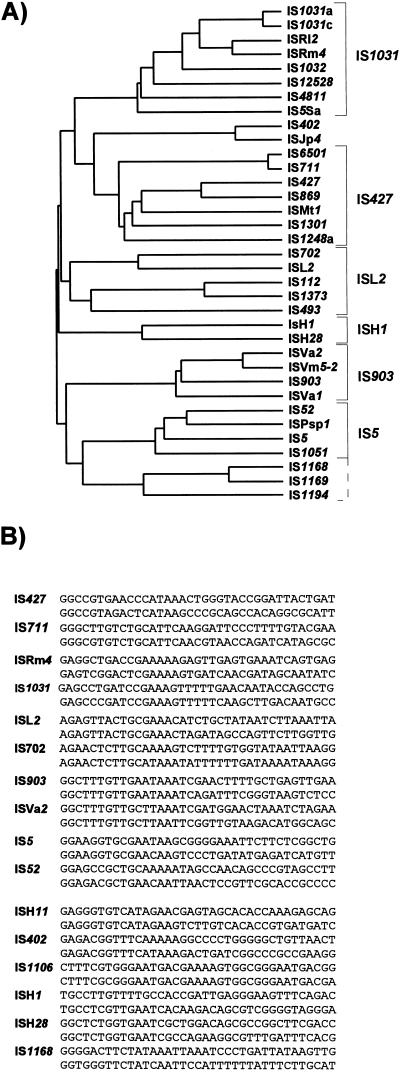

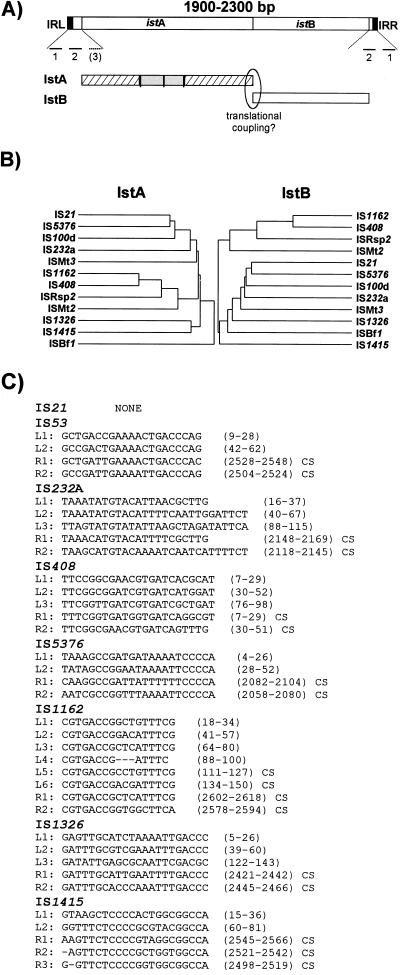
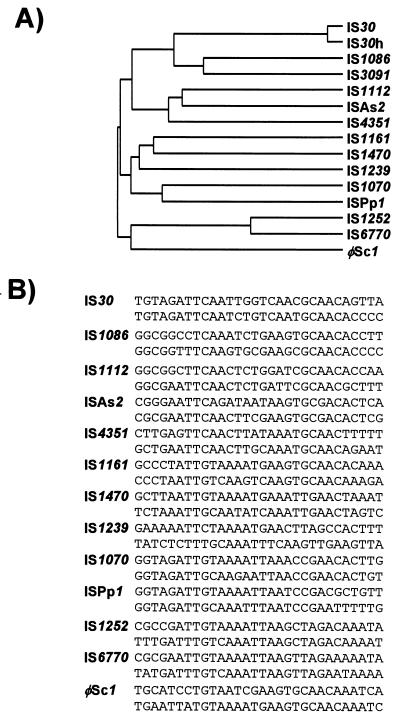
References
-
- Reference deleted.
-
- Abremski K E, Hoess R H. Evidence for a second conserved arginine residue in the integrase family of recombination proteins. Protein Eng. 1992;5:87–91. - PubMed
-
- Adzuma K, Mizuuchi K. Target immunity of Mu transposition reflects a differential distribution of Mu B protein. Cell. 1988;53:257–266. - PubMed
-
- Adzuma K, Mizuuchi K. Interaction of proteins located at a distance along DNA: mechanism of target immunity in the Mu DNA strand-transfer reaction. Cell. 1989;57:41–47. - PubMed
-
- Adzuma K, Mizuuchi K. Steady-state kinetic analysis of ATP hydrolysis by the B protein of bacteriophage Mu. Involvement of protein oligomerization in the ATPase cycle. J Biol Chem. 1991;266:6159–6167. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
