

CLB5 and CLB6 are required for premeiotic DNA replication and activation of the meiotic S/M checkpoint
- PMID: 9732268
- PMCID: PMC317137
- DOI: 10.1101/gad.12.17.2698
CLB5 and CLB6 are required for premeiotic DNA replication and activation of the meiotic S/M checkpoint
Abstract
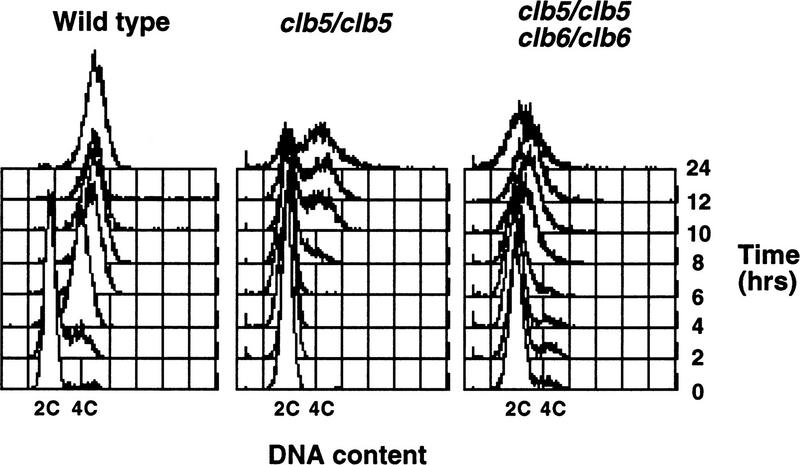
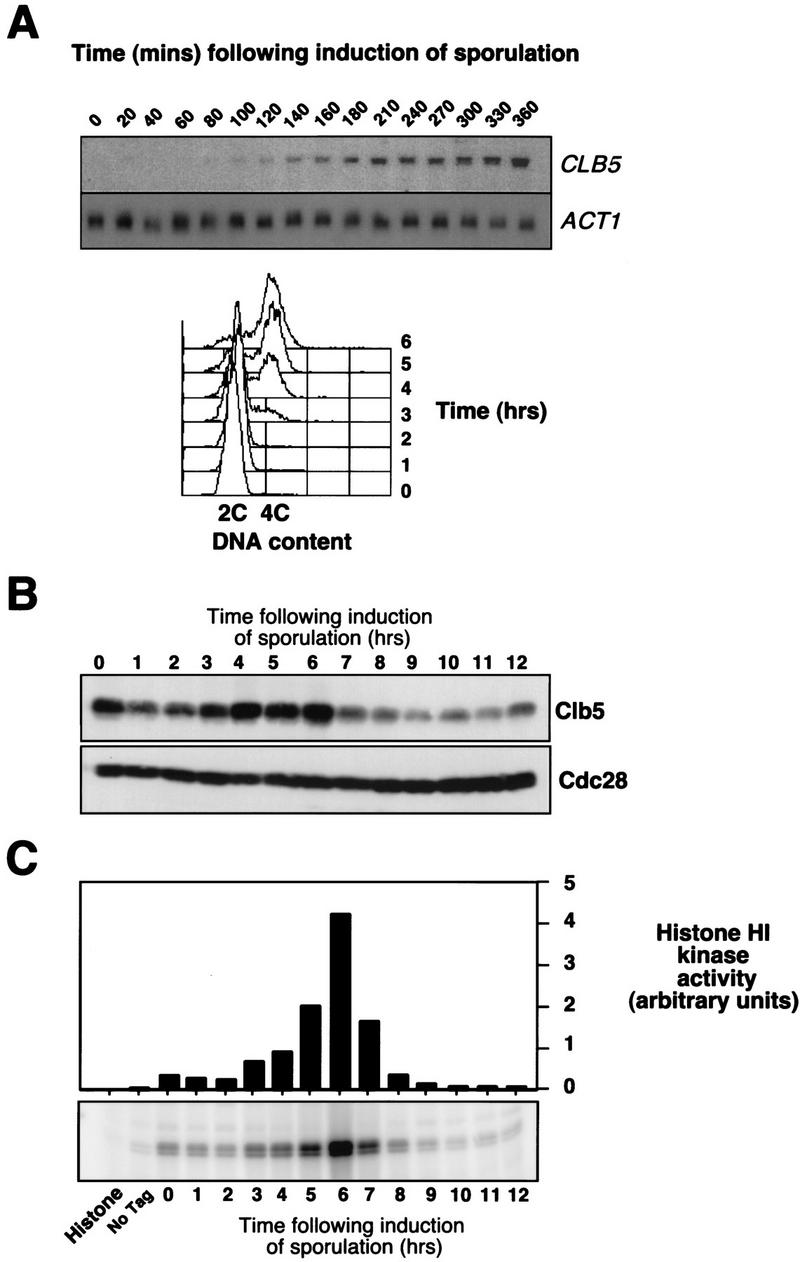
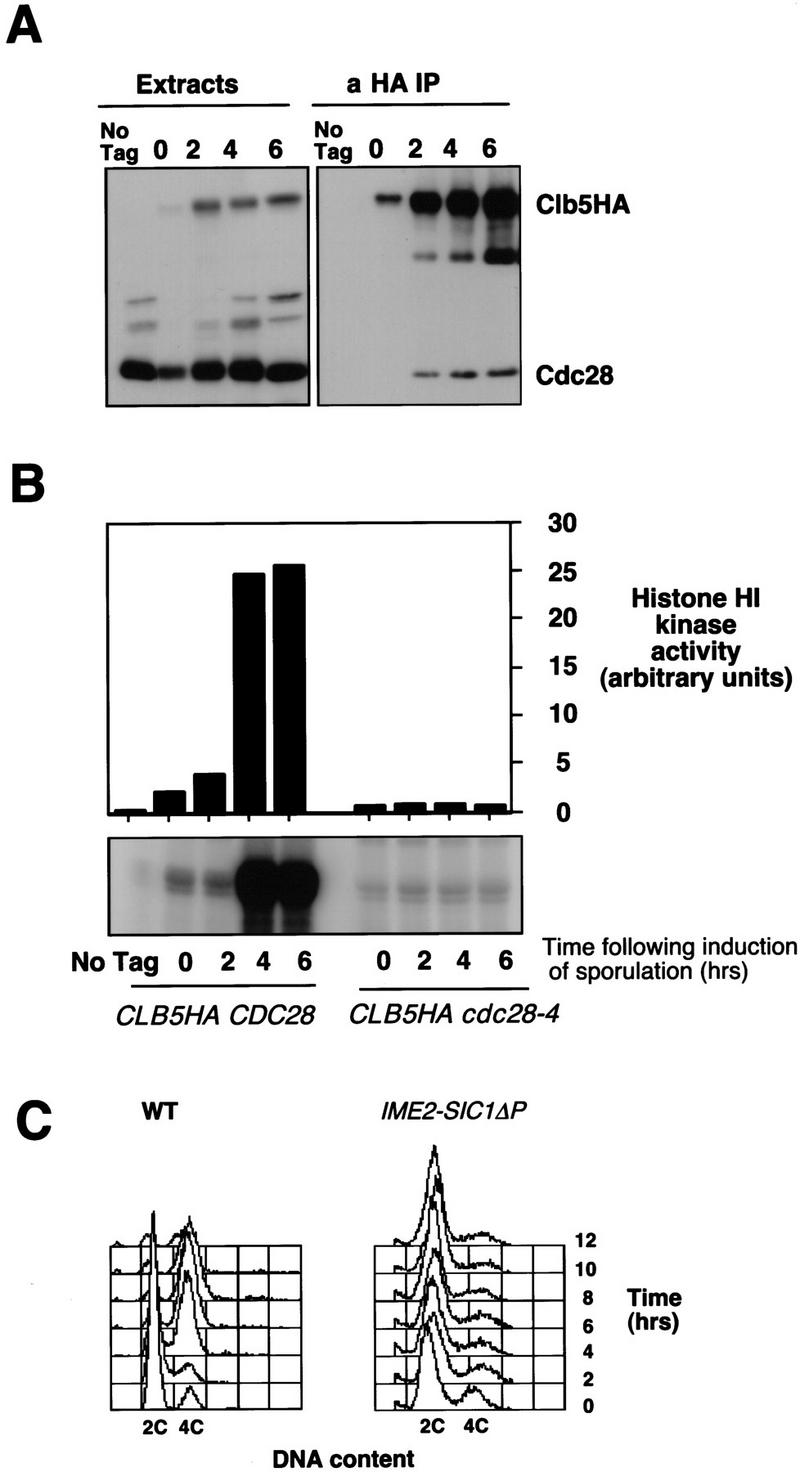
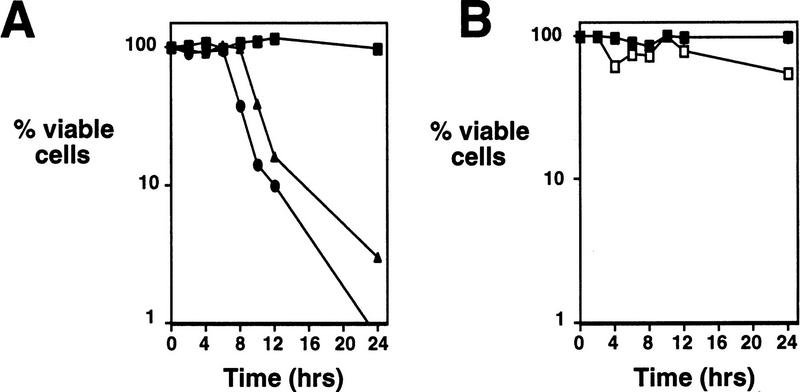
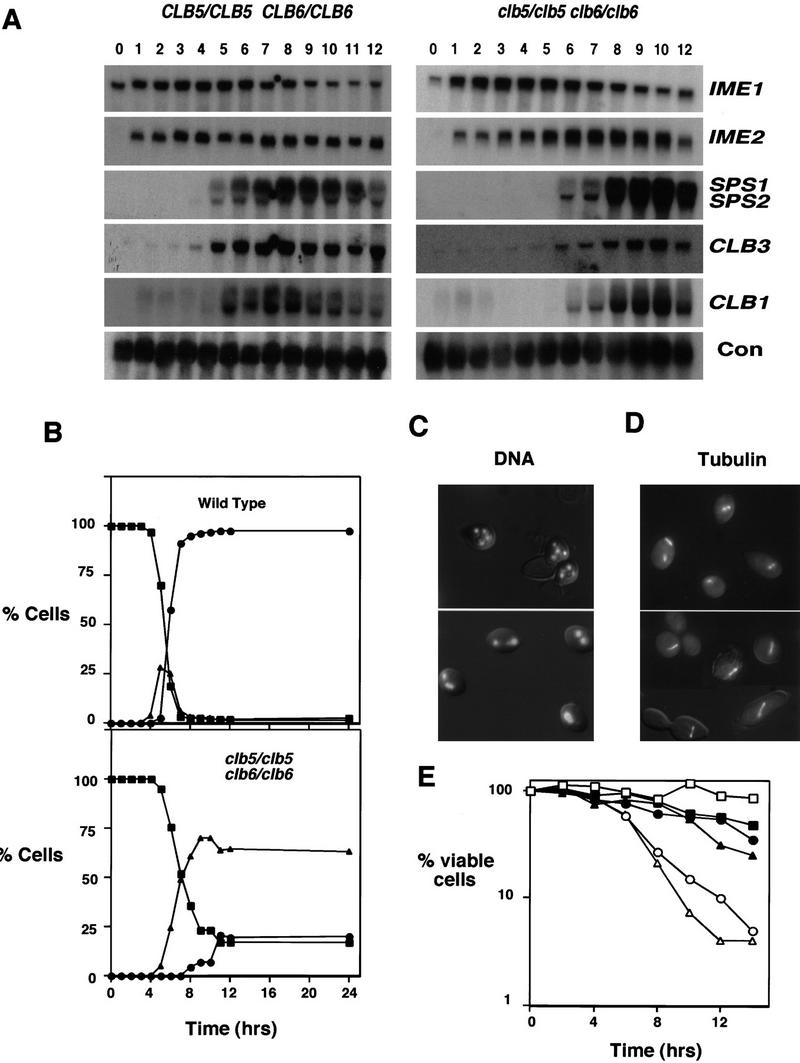
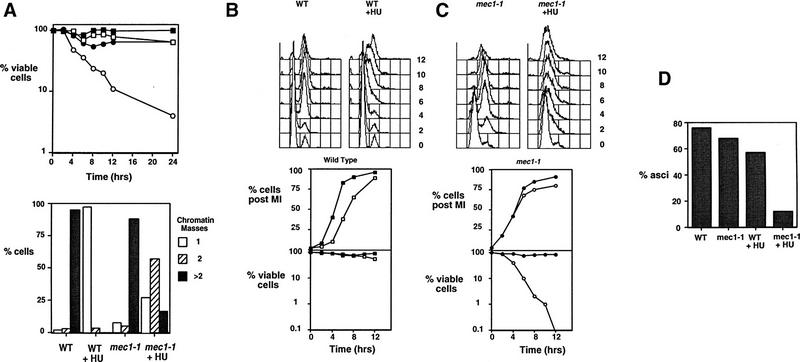
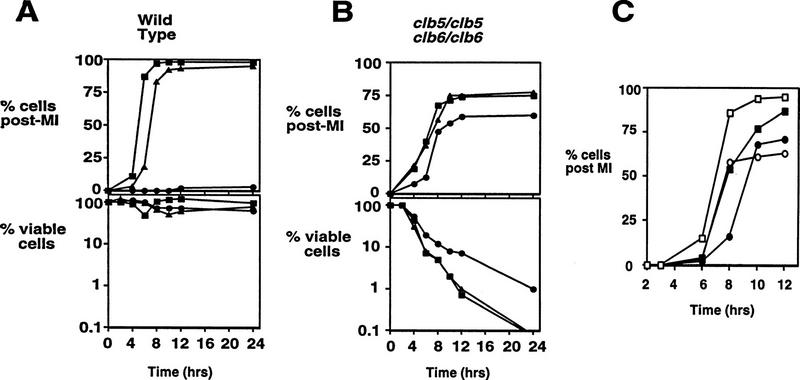
Initiation of DNA replication during the mitotic cell cycle requires the activation of a cyclin-dependent protein kinase (CDK). The B-type cyclins Clb5 and Clb6 are the primary activators of the S phase function of the budding yeast CDK Cdc28. However, in mitotically growing cells this role can be fulfilled by the other B-type cyclins Clb1-Clb4. We report here that cells undergoing meiotic development also require Clb dependent CDK activity for DNA replication. Diploid clb5/clb5 clb6/clb6 mutants are unable to perform premeiotic DNA replication. Despite this defect, the mutant cells progress into the meiotic program and undergo lethal segregation of unreplicated DNA suggesting that they fail to activate a checkpoint that restrains meiotic M phase until DNA replication is complete. We have found that a DNA replication checkpoint dependent on the ATM homolog MEC1 operates in wild-type cells during meiosis and can be invoked in response to inhibition of DNA synthesis. Although cells that lack clb5 and clb6 are unable to activate the meiotic DNA replication checkpoint, they do possess an intact DNA damage checkpoint which can restrain chromosome segregation in the face of DNA damage. We conclude that CLB5 and CLB6 are essential for premeiotic DNA replication and, consequently, for activation of a meiotic DNA replication checkpoint.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous