

Degradation of misfolded endoplasmic reticulum glycoproteins in Saccharomyces cerevisiae is determined by a specific oligosaccharide structure
- PMID: 9732283
- PMCID: PMC2149342
- DOI: 10.1083/jcb.142.5.1223
Degradation of misfolded endoplasmic reticulum glycoproteins in Saccharomyces cerevisiae is determined by a specific oligosaccharide structure
Abstract
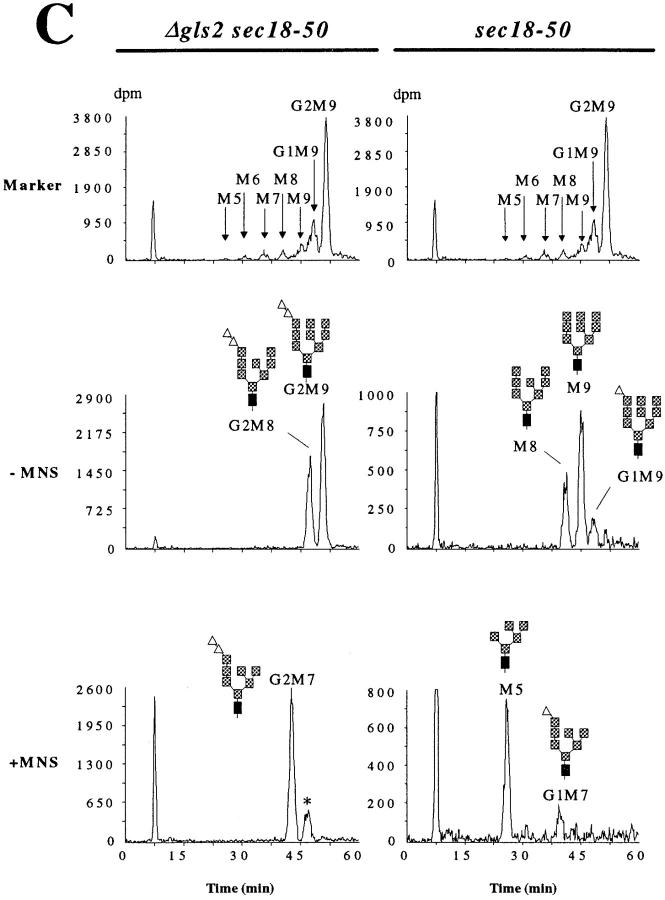
In Saccharomyces cerevisiae, transfer of N-linked oligosaccharides is immediately followed by trimming of ER-localized glycosidases. We analyzed the influence of specific oligosaccharide structures for degradation of misfolded carboxypeptidase Y (CPY). By studying the trimming reactions in vivo, we found that removal of the terminal alpha1,2 glucose and the first alpha1,3 glucose by glucosidase I and glucosidase II respectively, occurred rapidly, whereas mannose cleavage by mannosidase I was slow. Transport and maturation of correctly folded CPY was not dependent on oligosaccharide structure. However, degradation of misfolded CPY was dependent on specific trimming steps. Degradation of misfolded CPY with N-linked oligosaccharides containing glucose residues was less efficient compared with misfolded CPY bearing the correctly trimmed Man8GlcNAc2 oligosaccharide. Reduced rate of degradation was mainly observed for misfolded CPY bearing Man6GlcNAc2, Man7GlcNAc2 and Man9GlcNAc2 oligosaccharides, whereas Man8GlcNAc2 and, to a lesser extent, Man5GlcNAc2 oligosaccharides supported degradation. These results suggest a role for the Man8GlcNAc2 oligosaccharide in the degradation process. They may indicate the presence of a Man8GlcNAc2-binding lectin involved in targeting of misfolded glycoproteins to degradation in S. cerevisiae.
Figures







References
-
- Aebi M, Gassenhuber J, Domdey H, te Heesen S. Cloning and characterization of the ALG3 gene of Saccharomyces cerevisiae. . Glycobiology. 1996;6:439–444. - PubMed
-
- Alonso, J.M., A. Santa-Cecilia, and P. Calvo. Effect of bromoconduritol on glucosidase II from rat liver. Eur. J. Biochem. 215:37–42. - PubMed
-
- Bischoff J, Kornfeld R. Evidence for an α-mannosidase in endoplasmic reticulum of rat liver. J Biol Chem. 1983;258:7907–7910. - PubMed
-
- Bonifacino JS. Reversal of fortune for nascent proteins. Nature. 1996;384:405–406. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
