

Chilling delays circadian pattern of sucrose phosphate synthase and nitrate reductase activity in tomato
- PMID: 9733534
- PMCID: PMC34851
- DOI: 10.1104/pp.118.1.149
Chilling delays circadian pattern of sucrose phosphate synthase and nitrate reductase activity in tomato
Abstract
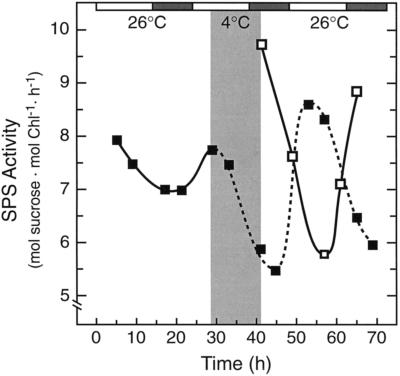
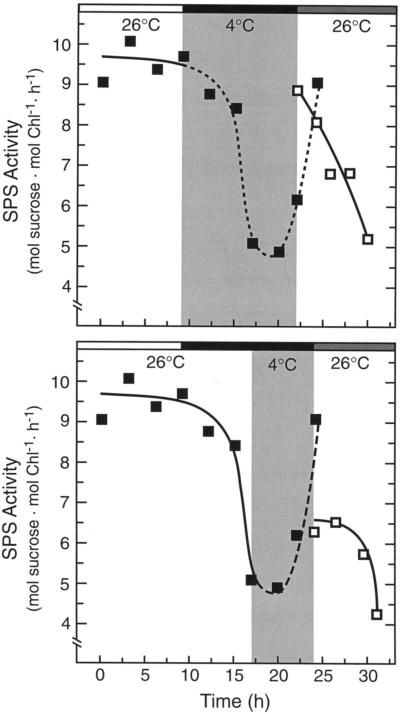
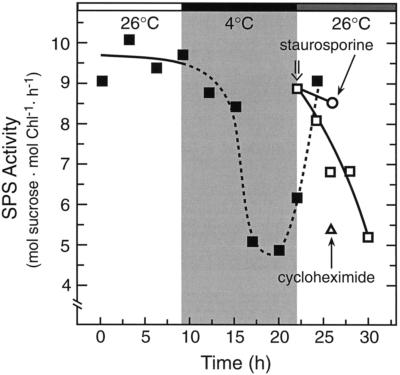
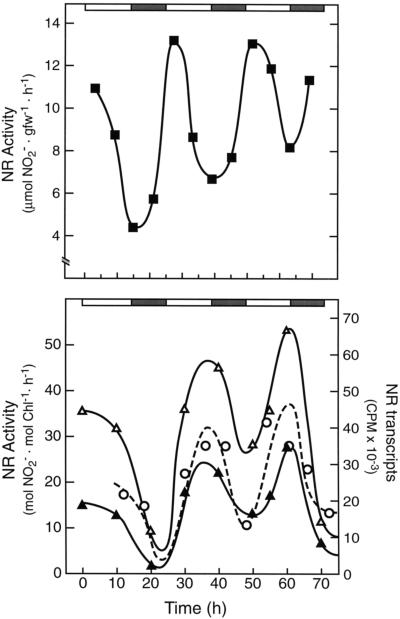
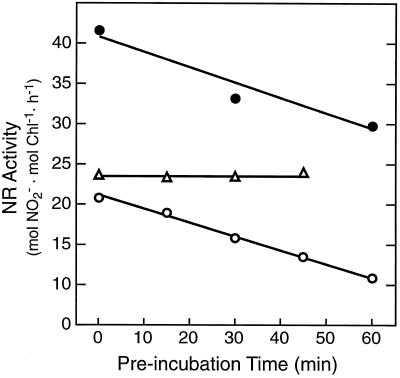
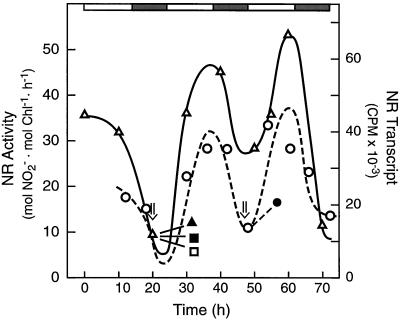
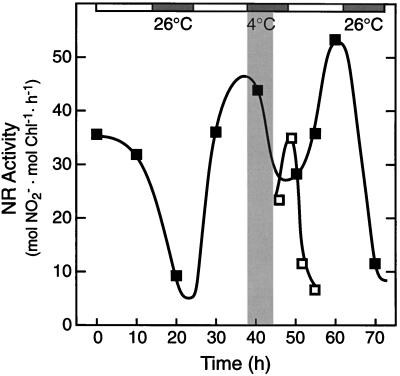
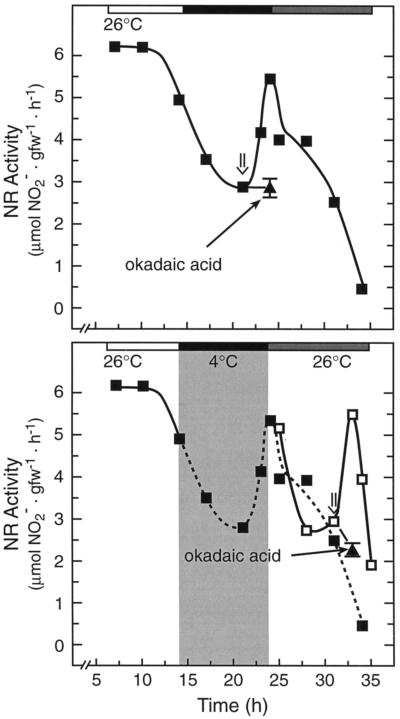
Overnight low-temperature exposure inhibits photosynthesis in chilling-sensitive species such as tomato (Lycopersicon esculentum) and cucumber by as much as 60%. In an earlier study we showed that one intriguing effect of low temperature on chilling-sensitive plants is to stall the endogenous rhythm controlling transcription of certain nuclear-encoded genes, causing the synthesis of the corresponding transcripts and proteins to be mistimed when the plant is rewarmed. Here we show that the circadian rhythm controlling the activity of sucrose phosphate synthase (SPS) and nitrate reductase (NR), key control points of carbon and nitrogen metabolism in plant cells, is delayed in tomato by chilling treatments. Using specific protein kinase and phosphatase inhibitors, we further demonstrate that the chilling-induced delay in the circadian control of SPS and NR activity is associated with the activity of critical protein phosphatases. The sensitivity of the pattern of SPS activity to specific inhibitors of transcription and translation indicates that there is a chilling-induced delay in SPS phosphorylation status that is caused by an effect of low temperature on the expression of a gene coding for a phosphoprotein phosphatase, perhaps the SPS phosphatase. In contrast, the chilling-induced delay in NR activity does not appear to arise from effects on NR phosphorylation status, but rather from direct effects on NR expression. It is likely that the mistiming in the regulation of SPS and NR, and perhaps other key metabolic enzymes under circadian regulation, underlies the chilling sensitivity of photosynthesis in these plant species.
Figures
References
-
- Baker NR, East TM, Long SP. Chilling damage to photosynthesis in young Zea mays. II. Photochemical function of thylakoids in vivo. J Exp Bot. 1983;34:189–197.
-
- Bringloe DH, Dyer TA, Gray JC. Developmental, circadian and light regulation of wheat ferredoxin gene expression. Plant Mol Biol. 1995;27:293–306. - PubMed
LinkOut - more resources
Full Text Sources
