

The biosynthesis of erucic acid in developing embryos of brassica rapa
- PMID: 9733537
- PMCID: PMC34854
- DOI: 10.1104/pp.118.1.183
The biosynthesis of erucic acid in developing embryos of brassica rapa
Abstract
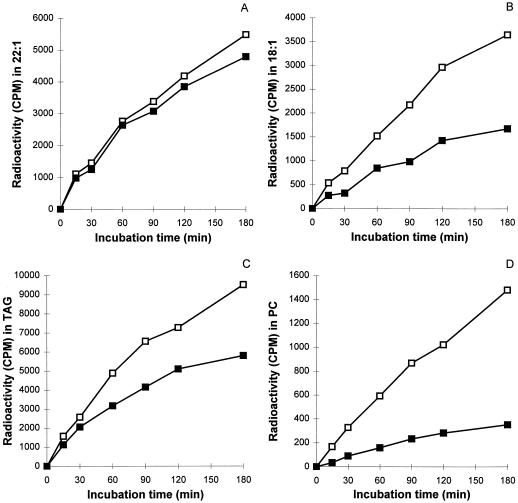
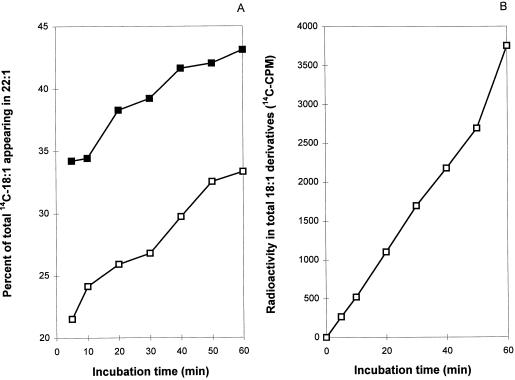
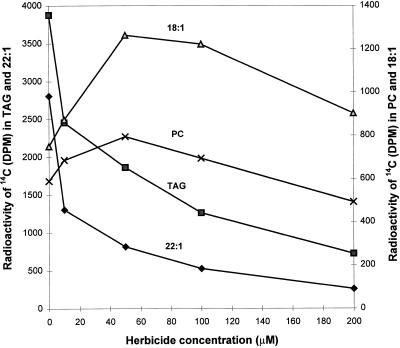
The prevailing hypothesis on the biosynthesis of erucic acid in developing seeds is that oleic acid, produced in the plastid, is activated to oleoyl-coenzyme A (CoA) for malonyl-CoA-dependent elongation to erucic acid in the cytosol. Several in vivo-labeling experiments designed to probe and extend this hypothesis are reported here. To examine whether newly synthesized oleic acid is directly elongated to erucic acid in developing seeds of Brassica rapa L., embryos were labeled with [14C]acetate, and the ratio of radioactivity of carbon atoms C-5 to C-22 (de novo fatty acid synthesis portion) to carbon atoms C-1 to C-4 (elongated portion) of erucic acid was monitored with time. If newly synthesized 18:1 (oleate) immediately becomes a substrate for elongation to erucic acid, this ratio would be expected to remain constant with incubation time. However, if erucic acid is produced from a pool of preexisting oleic acid, the ratio of 14C in the 4 elongation carbons to 14C in the methyl-terminal 18 carbons would be expected to decrease with time. This labeling ratio decreased with time and, therefore, suggests the existence of an intermediate pool of 18:1, which contributes at least part of the oleoyl precursor for the production of erucic acid. The addition of 2-[3-chloro-5-(trifluromethyl)-2-pyridinyloxyphenoxy] propanoic acid, which inhibits the homodimeric acetyl-CoA carboxylase, severely inhibited the synthesis of [14C]erucic acid, indicating that essentially all malonyl-CoA for elongation of 18:1 to erucate was produced by homodimeric acetyl-CoA carboxylase. Both light and 2-[3-chloro-5-(trifluromethyl)-2-pyridinyloxyphenoxy]-propanoic acid increased the accumulation of [14C]18:1 and the parallel accumulation of [14C]phosphatidylcholine. Taken together, these results show an additional level of complexity in the biosynthesis of erucic acid.
Figures

References
-
- Appleby RS, Gurr MI, Nichols BW. Studies on seed-oil triglycerides: factor controlling biosynthesis of fatty acid and acyl lipids in subcellular organelles of maturing Crambe abyssinica seeds. Eur J Biochem. 1974;48:209–216. - PubMed
-
- Asokanthan PS, Johnson RW, Griffith M, Krol M. The photosynthetic potential of canola embryos. Physiol Plant. 1997;101:353–360.
-
- Bligh EG, Dyer WJ. A rapid method of total lipid extraction and purification. Can J Biochem Physiol. 1959;37:911–917. - PubMed
-
- Browse J, Slack CR. Fatty-acid synthesis in plastids from maturing safflower and linseed cotyledons. Planta. 1985;166:74–80. - PubMed
-
- Burton JD, Gronwald JW, Keith RA, Somers DA, Gengenbach BG, Wyse DL. Kinetics of inhibition of plant acetyl-coenzyme A carboxylase by sethoxydim and haloxyfop. Pestic Biochem Physiol. 1991;39:100–109.
LinkOut - more resources
Full Text Sources
Miscellaneous
