

Differential control of xanthophylls and light-induced stress proteins, as opposed to light-harvesting chlorophyll a/b proteins, during photosynthetic acclimation of barley leaves to light irradiance
- PMID: 9733542
- PMCID: PMC34860
- DOI: 10.1104/pp.118.1.227
Differential control of xanthophylls and light-induced stress proteins, as opposed to light-harvesting chlorophyll a/b proteins, during photosynthetic acclimation of barley leaves to light irradiance
Abstract
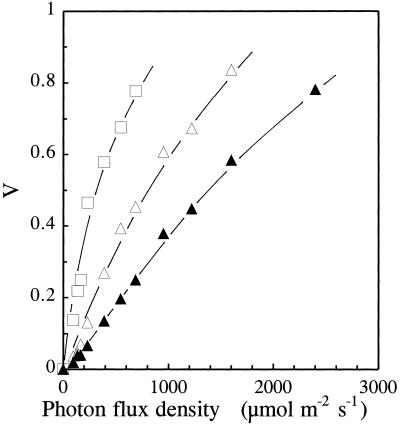
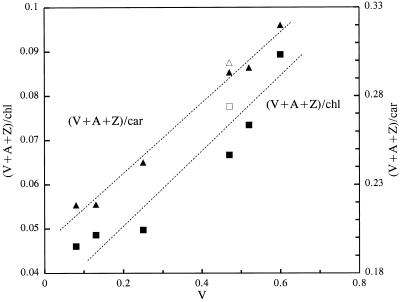
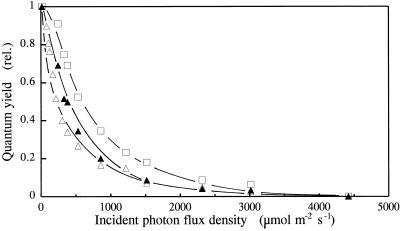
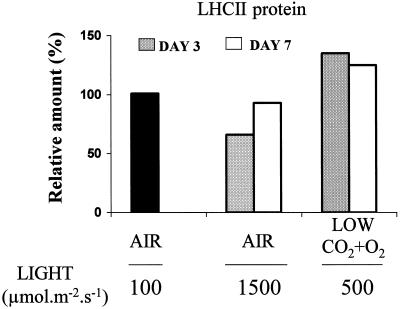
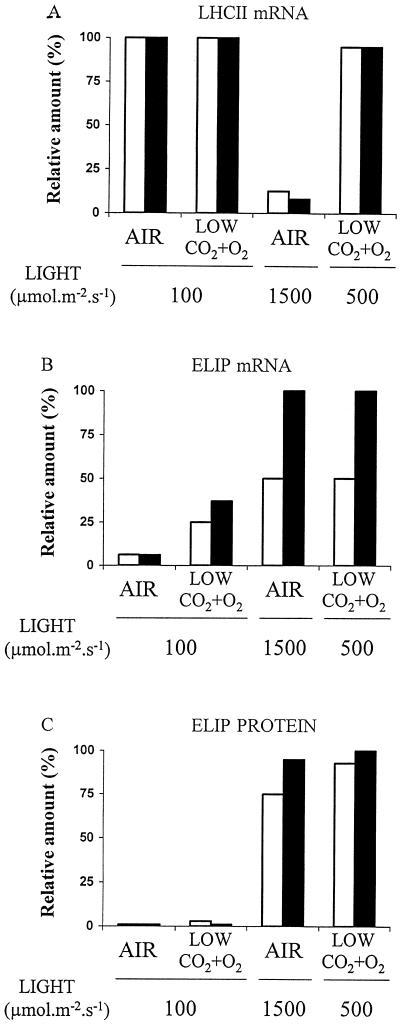
Barley (Hordeum vulgare L.) plants were grown at different photon flux densities ranging from 100 to 1800 &mgr;mol m-2 s-1 in air and/or in atmospheres with reduced levels of O2 and CO2. Low O2 and CO2 partial pressures allowed plants to grow under high photosystem II (PSII) excitation pressure, estimated in vivo by chlorophyll fluorescence measurements, at moderate photon flux densities. The xanthophyll-cycle pigments, the early light-inducible proteins, and their mRNA accumulated with increasing PSII excitation pressure irrespective of the way high excitation pressure was obtained (high-light irradiance or decreased CO2 and O2 availability). These findings indicate that the reduction state of electron transport chain components could be involved in light sensing for the regulation of nuclear-encoded chloroplast gene expression. In contrast, no correlation was found between the reduction state of PSII and various indicators of the PSII light-harvesting system, such as the chlorophyll a-to-b ratio, the abundance of the major pigment-protein complex of PSII (LHCII), the mRNA level of LHCII, the light-saturation curve of O2 evolution, and the induced chlorophyll-fluorescence rise. We conclude that the chlorophyll antenna size of PSII is not governed by the redox state of PSII in higher plants and, consequently, regulation of early light-inducible protein synthesis is different from that of LHCII.
Figures
References
-
- Adams WW, III, Demmig-Adams B, Verhoeven AS, Barker DH. ‘Photoinhibition’ during winter stress: involvement of sustained xanthophyll cycle-dependent energy dissipation. Aust J Plant Physiol. 1994;22:261–276.
-
- Adamska I. ELIPs: light-induced stress proteins. Physiol Plant. 1997;100:794–805.
-
- Adamska I, Kloppstech K. Low temperature increases the abundance of early light-inducible transcript under light stress conditions. J Biol Chem. 1994;269:30221–30226. - PubMed
-
- Adamska I, Kloppstech K, Ohad I. The effect of free radical enhancers and scavengers on accumulation of early light-inducible protein during light stress. Z Naturforsch. 1993;48c:391–396.
LinkOut - more resources
Full Text Sources
