

trans-encapsidation of a poliovirus replicon by different picornavirus capsid proteins
- PMID: 9733835
- PMCID: PMC110132
- DOI: 10.1128/JVI.72.10.7972-7977.1998
trans-encapsidation of a poliovirus replicon by different picornavirus capsid proteins
Abstract
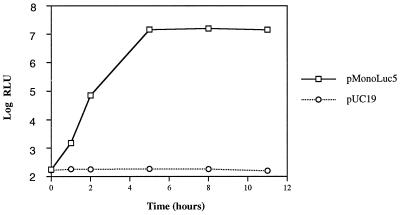
A trans-encapsidation assay was established to study the specificity of picornavirus RNA encapsidation. A poliovirus replicon with the luciferase gene replacing the capsid protein-coding region was coexpressed in transfected HeLa cells with capsid proteins from homologous or heterologous virus. Successful trans-encapsidation resulted in assembly and production of virions whose replication, upon subsequent infection of HeLa cells, was accompanied by expression of luciferase activity. The amount of luciferase activity was proportional to the amount of trans-encapsidated virus produced from the cotransfection. When poliovirus capsid proteins were supplied in trans, >2 x 10(6) infectious particles/ml were produced. When coxsackievirus B3, human rhinovirus 14, mengovirus, or hepatitis A virus (HAV) capsid proteins were supplied in trans, all but HAV showed some encapsidation of the replicon. The overall encapsidation efficiency of the replicon RNA by heterologous capsid proteins was significantly lower than when poliovirus capsid was used. trans-encapsidated particles could be completely neutralized with specific antisera against each of the donor virus capsids. The results indicate that encapsidation is regulated by specific viral nucleic acid and protein sequences.
Figures
Similar articles
-
Encapsidation of poliovirus replicons encoding the complete human immunodeficiency virus type 1 gag gene by using a complementation system which provides the P1 capsid protein in trans.J Virol. 1995 Mar;69(3):1548-55. doi: 10.1128/JVI.69.3.1548-1555.1995. J Virol. 1995. PMID: 7853488 Free PMC article.
-
Demonstration of the specificity of poliovirus encapsidation using a novel replicon which encodes enzymatically active firefly luciferase.Virology. 1998 Mar 30;243(1):1-11. doi: 10.1006/viro.1998.9046. Virology. 1998. PMID: 9527910
-
Inherent instability of poliovirus genomes containing two internal ribosome entry site (IRES) elements supports a role for the IRES in encapsidation.J Virol. 2000 Sep;74(18):8335-42. doi: 10.1128/jvi.74.18.8335-8342.2000. J Virol. 2000. PMID: 10954532 Free PMC article.
-
Picornavirus morphogenesis.Microbiol Mol Biol Rev. 2014 Sep;78(3):418-37. doi: 10.1128/MMBR.00012-14. Microbiol Mol Biol Rev. 2014. PMID: 25184560 Free PMC article. Review.
-
Evolutionary and Structural Overview of Human Picornavirus Capsid Antibody Evasion.Front Cell Infect Microbiol. 2019 Aug 20;9:283. doi: 10.3389/fcimb.2019.00283. eCollection 2019. Front Cell Infect Microbiol. 2019. PMID: 31482072 Free PMC article. Review.
Cited by
-
Trans-Encapsidation of Foot-and-Mouth Disease Virus Genomes Facilitates Escape from Neutralizing Antibodies.Viruses. 2022 May 27;14(6):1161. doi: 10.3390/v14061161. Viruses. 2022. PMID: 35746633 Free PMC article.
-
Identification of a cis-acting replication element within the poliovirus coding region.J Virol. 2000 May;74(10):4590-600. doi: 10.1128/jvi.74.10.4590-4600.2000. J Virol. 2000. PMID: 10775595 Free PMC article.
-
Coxsackievirus expression of the murine secretory protein interleukin-4 induces increased synthesis of immunoglobulin G1 in mice.J Virol. 2000 Sep;74(17):7952-62. doi: 10.1128/jvi.74.17.7952-7962.2000. J Virol. 2000. PMID: 10933703 Free PMC article.
-
Formation of the poliovirus replication complex requires coupled viral translation, vesicle production, and viral RNA synthesis.J Virol. 2000 Jul;74(14):6570-80. doi: 10.1128/jvi.74.14.6570-6580.2000. J Virol. 2000. PMID: 10864671 Free PMC article.
-
Direct interaction between two viral proteins, the nonstructural protein 2C and the capsid protein VP3, is required for enterovirus morphogenesis.PLoS Pathog. 2010 Aug 26;6(8):e1001066. doi: 10.1371/journal.ppat.1001066. PLoS Pathog. 2010. PMID: 20865167 Free PMC article.
References
-
- Andino R, Silvera D, Suggett S D, Achacoso P L, Miller C J, Baltimore D, Feinberg M B. Engineering poliovirus as a vaccine vector for the expression of diverse antigens. Science. 1994;265:1448–1451. - PubMed
-
- Baltimore D. The replication of picornaviruses. In: Levy H B, editor. The biochemistry of viruses. New York, N.Y: Marcel Dekker; 1969. pp. 101–176.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
