

The RNA polymerase of influenza virus, bound to the 5' end of virion RNA, acts in cis to polyadenylate mRNA
- PMID: 9733864
- PMCID: PMC110172
- DOI: 10.1128/JVI.72.10.8214-8219.1998
The RNA polymerase of influenza virus, bound to the 5' end of virion RNA, acts in cis to polyadenylate mRNA
Abstract
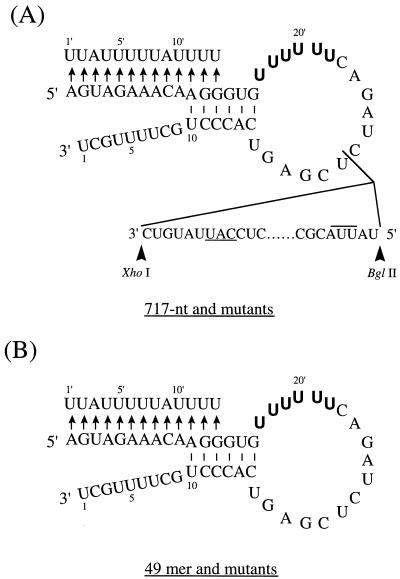
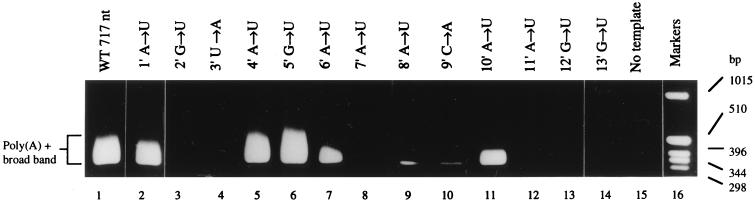
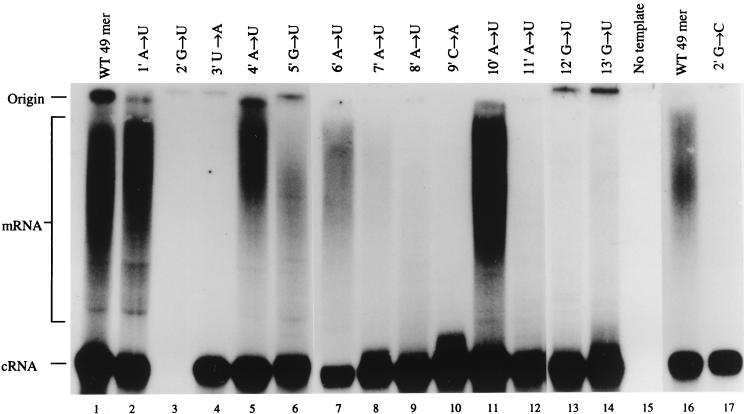
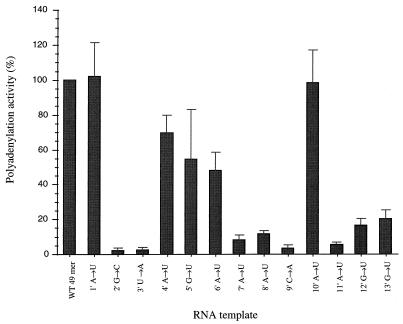
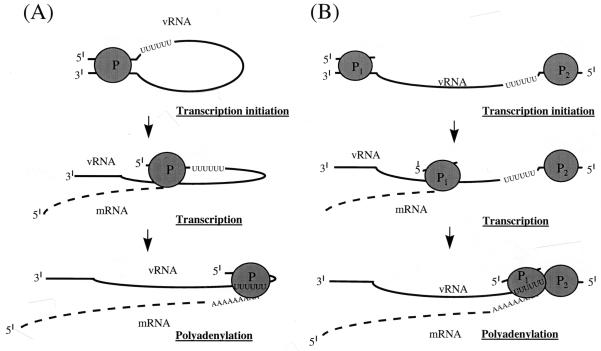
We previously demonstrated, by limited mutagenesis, that conserved sequence elements within the 5' end of influenza virus virion RNA (vRNA) are required for the polyadenylation of mRNA in vitro. To further characterize the nucleotide residues at the 5' end of vRNA which might be involved in polyadenylation, a complete set of short and long model vRNA-like templates with mutations at nucleotides 1' to 13' (prime notation denotes numbering from the 5' end) of vRNA were synthesized and transcribed in vitro. The products were assayed for mRNA production with both reverse transcription-PCR and [alpha-32P]ATP incorporation assays. Results from these independent assays showed that vRNA templates with point mutations at positions 2', 3', 7' to 9', and 11' to 13' synthesized polyadenylated transcripts inefficiently compared with those with mutations at positions 1', 4' to 6', and 10'. Positions 2', 3', 7' to 9', and 11' are known to be involved in RNA polymerase binding. Furthermore, residues at positions 11' to 13' are known to be involved in base pairing between the 3' and 5' ends of vRNA. These findings demonstrate that the RNA polymerase has to bind to the 5' end of the template vRNA, which must then interact with the 3' end of the same template for polyadenylation to occur. These results support a model in which a cis-acting RNA polymerase is required for the polyadenylation of influenza virus.
Figures
References
-
- Desselberger U, Racaniello V R, Zazra J J, Palese P. The 3′ and 5′ terminal sequences of influenza A, B and C virus RNA segments are highly conserved and show partial inverted complementarity. Gene. 1980;8:315–328. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
