

doi: 10.1128/JVI.72.10.8349-8353.1998.
Monitoring retroviral RNA dimerization in vivo via hammerhead ribozyme cleavage
Affiliations
- PMID: 9733882
- PMCID: PMC110209
- DOI: 10.1128/JVI.72.10.8349-8353.1998
Item in Clipboard
Monitoring retroviral RNA dimerization in vivo via hammerhead ribozyme cleavage
J Virol.
1998 Oct.
Abstract
We have used a strategy for colocalization of Psi (Psi)-tethered ribozymes and targets to demonstrate that Psi sequences are capable of specific interaction in the cytoplasm of both packaging and nonpackaging cells. These results indicate that current in vitro dimerization models may have in vivo counterparts. The methodology used may be applied to further genetic analyses on Psi domain interactions in vivo.
Figures
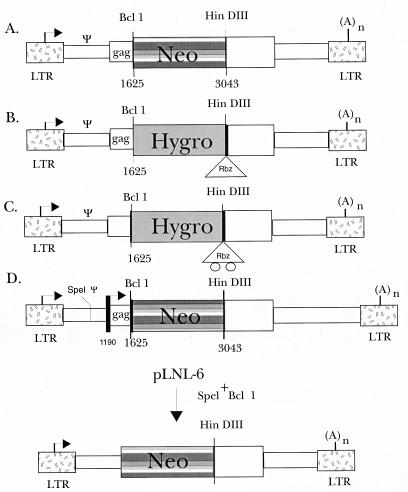
Linear maps of retroviral vectors used in this study. The MoMuLV-based retroviral vector, pLNL-6 (pΨneo) was kindly provided by A. Dusty Miller of the Fred Hutchinson Cancer Research Center, Seattle, Wash. (1), and served as the basic vector for all of the constructs in this study. Target plasmids pΨneo (A) and pΔΨneo (D) contain the neomycin phosphotransferase (neo) gene, allowing selection of transfected cells with G418. Retroviral vectors expressing a ribozyme (pΨhygRbz) (B) or a mutant ribozyme (pΨhyg*Rbz) (C) were derived from pLNL-6 by replacement of the neo gene with hygromycin phosphotransferase (hygro)-Rbz or hygro-*Rbz constructs. Specifically, a 1.1-kb BamHI fragment containing the hygro gene was subcloned upstream of either a functional or a mutant ribozyme gene in pBluescript. The hygro-ribozyme segments were inserted into the BclI and HindIII sites of pLNL-6 (B and C). This functionally destroyed both the BclI and BamHI sites in the LTR-hygromycin-LTR (LHL) vectors.
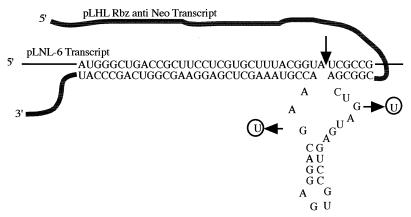
Pairing of the anti-neo ribozyme with its target in LNL-6. The ribozyme cleavage site in the neo transcript is between nucleotides 2391 and 2392 in the pLNL-6 sequence and is approximately 1,800 nucleotides downstream of the start point of transcription. Since the ribozyme cloning site in the pΨhygRbz vector was constrained to a site just downstream of the hygro coding region, the neo target site was chosen to position it at the same relative distance from the messenger cap site as the ribozyme, to enhance the probability of the ribozyme pairing arms interacting with the target sequence in the dimerized RNAs. The ribozyme has asymmetric base pairing, with 6 bp in stem I and 32 bp in stem III. The long stem III facilitates the ribozyme and target association, whereas once cleavage has taken place, the short stem I facilitates rapid product dissociation and possibly hastens the ultimate degradation of the cleaved RNA (16). To control for ribozyme versus antisense activity, we generated a mutant, inactive version of the anti-neo ribozyme and inserted it in place of the functional ribozyme in pΨhyg*Rbz (Fig. 1C).
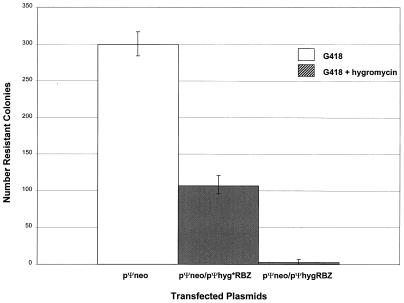
Ribozyme-mediated inhibition of G418-resistant transfectants in cultured PA317 Cells. The columns show the average numbers of transformed colonies obtained after G418 plus hygromycin selection from three separate experiments; the error bars indicate the data range.
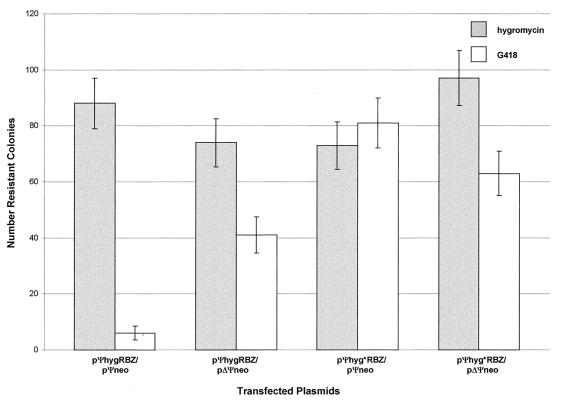
Ψ-mediated colocalization of ribozyme and target RNAs in PA317 packaging cells. Each column shows the average number of colonies for duplicate transfections obtained under the indicated selection conditions; the error bars show the data range.
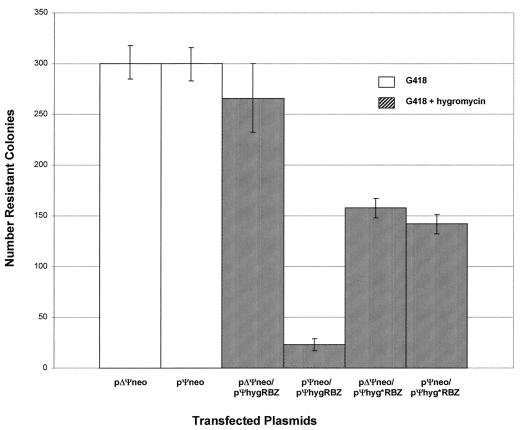
Ribozyme activity assay of Ψ-mediated colocalization in PA317 packaging cells. In the negative control (no plasmid), all cells died within 1 week. Each column shows the average number of colonies for duplicate transfections obtained under the indicated selection conditions; the error bars show the data range.
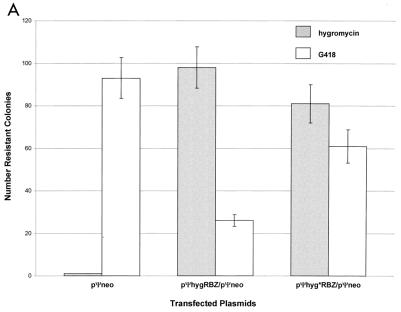
Ψ-mediated colocalization of RNAs in 293 nonpackaging cells. Each column shows the average number of colonies for duplicate transfections obtained under the indicated selection conditions; the error bars show the data range. (A) Ribozyme-specific inhibition of target expression; (B) Ψ-dependent inhibition of target expression.
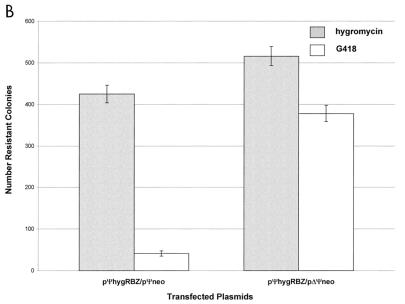
Ψ-mediated colocalization of RNAs in 293 nonpackaging cells. Each column shows the average number of colonies for duplicate transfections obtained under the indicated selection conditions; the error bars show the data range. (A) Ribozyme-specific inhibition of target expression; (B) Ψ-dependent inhibition of target expression.
Similar articles
-
Ribozyme-mediated suppression of Moloney murine leukemia virus and human immunodeficiency virus type I replication in permissive cell lines.Proc Natl Acad Sci U S A. 1994 Oct 11;91(21):9715-9. doi: 10.1073/pnas.91.21.9715. Proc Natl Acad Sci U S A. 1994. PMID: 7937878 Free PMC article.
-
Analytical study of rat retrotransposon VL30 RNA dimerization in vitro and packaging in murine leukemia virus.J Mol Biol. 1994 Jul 29;240(5):434-44. doi: 10.1006/jmbi.1994.1459. J Mol Biol. 1994. PMID: 8046749
-
Tethering ribozymes to a retroviral packaging signal for destruction of viral RNA.Science. 1993 Dec 3;262(5139):1566-9. doi: 10.1126/science.8248806. Science. 1993. PMID: 8248806
-
Ribozymes: structure and mechanism in RNA catalysis.Trends Biochem Sci. 1996 Jun;21(6):220-4. Trends Biochem Sci. 1996. PMID: 8744356 Review.
-
Ribozymes of the hepatitis delta virus: recent findings on their structure, mechanism of catalysis and possible applications.Acta Biochim Pol. 2001;48(2):409-18. Acta Biochim Pol. 2001. PMID: 11732611 Review.
Cited by
-
mRNA localization signals can enhance the intracellular effectiveness of hammerhead ribozymes.RNA. 1999 Sep;5(9):1200-9. doi: 10.1017/s1355838299990246. RNA. 1999. PMID: 10496221 Free PMC article.
-
Two distinct Moloney murine leukemia virus RNAs produced from a single locus dimerize at random.Virology. 2006 Jan 20;344(2):391-400. doi: 10.1016/j.virol.2005.09.002. Epub 2005 Oct 10. Virology. 2006. PMID: 16216294 Free PMC article.
-
Structure of an RNA switch that enforces stringent retroviral genomic RNA dimerization.Proc Natl Acad Sci U S A. 2006 Sep 12;103(37):13640-5. doi: 10.1073/pnas.0606156103. Epub 2006 Aug 31. Proc Natl Acad Sci U S A. 2006. PMID: 16945907 Free PMC article.
-
Nonrandom dimerization of murine leukemia virus genomic RNAs.J Virol. 2004 Nov;78(22):12129-39. doi: 10.1128/JVI.78.22.12129-12139.2004. J Virol. 2004. PMID: 15507599 Free PMC article.
References
-
- Alford R L, Honda S, Laurence C B, Belmont J W. RNA secondary structure analysis of the packaging signal for Moloney murine leukemia virus. Virology. 1991;183:611–619. - PubMed
-
- Awang G, Sen D. Mode of dimerization of HIV-1 genomic RNA. Biochemistry. 1993;32:11453–11457. - PubMed
-
- Bender W, Davidson N. Mapping of poly(A) sequences in the electron microscope reveals unusual structure of type C oncornavirus RNA molecules. Cell. 1976;7:595–607. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
