

The 2.8-A structure of rat liver F1-ATPase: configuration of a critical intermediate in ATP synthesis/hydrolysis
- PMID: 9736690
- PMCID: PMC21596
- DOI: 10.1073/pnas.95.19.11065
The 2.8-A structure of rat liver F1-ATPase: configuration of a critical intermediate in ATP synthesis/hydrolysis
Abstract
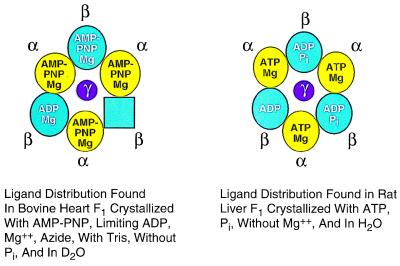
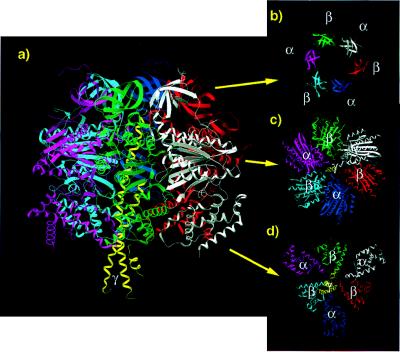
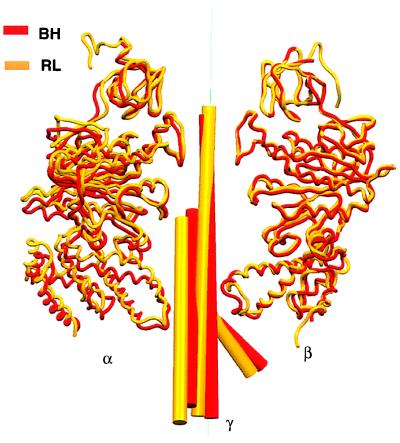
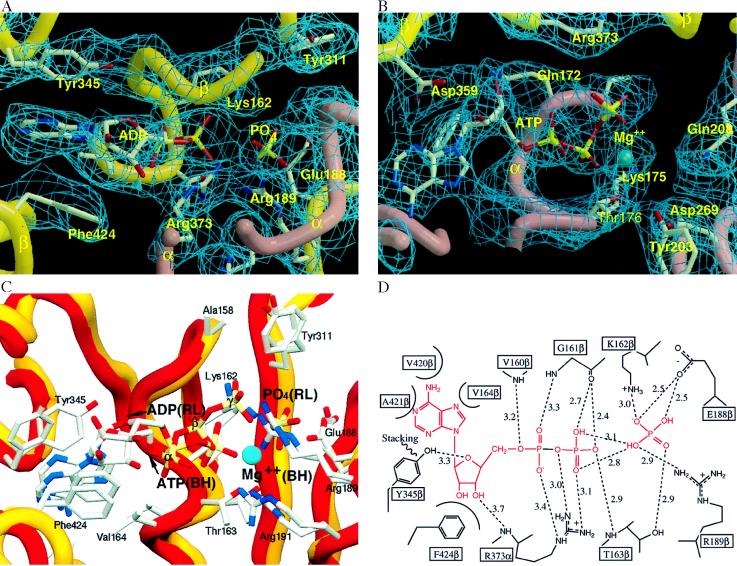
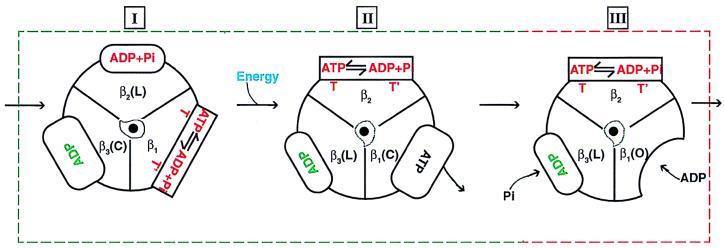
During mitochondrial ATP synthesis, F1-ATPase-the portion of the ATP synthase that contains the catalytic and regulatory nucleotide binding sites-undergoes a series of concerted conformational changes that couple proton translocation to the synthesis of the high levels of ATP required for cellular function. In the structure of the rat liver F1-ATPase, determined to 2.8-A resolution in the presence of physiological concentrations of nucleotides, all three beta subunits contain bound nucleotide and adopt similar conformations. This structure provides the missing configuration of F1 necessary to define all intermediates in the reaction pathway. Incorporation of this structure suggests a mechanism of ATP synthesis/hydrolysis in which configurations of the enzyme with three bound nucleotides play an essential role.
Figures
References
-
- Catterall W A, Pedersen P L. J Biol Chem. 1971;246:4987–4994. - PubMed
-
- Catterall W A, Coty W A, Pedersen P L. J Biol Chem. 1973;248:7427–7431. - PubMed
-
- Catterall W A, Pedersen P L. Biochem Soc Spec Publ. 1974;4:63–88.
-
- Boyer P D. Annu Rev Biochem. 1997;66:714–749. - PubMed
-
- Weber J, Senior A E. Biochim Biophys Acta. 1997;1319:19–58. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
