

An essential role for the phosphatidylinositol transfer protein in the scission of coatomer-coated vesicles from the trans-Golgi network
- PMID: 9736710
- PMCID: PMC21616
- DOI: 10.1073/pnas.95.19.11181
An essential role for the phosphatidylinositol transfer protein in the scission of coatomer-coated vesicles from the trans-Golgi network
Abstract
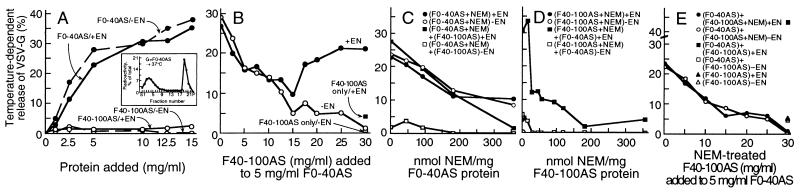
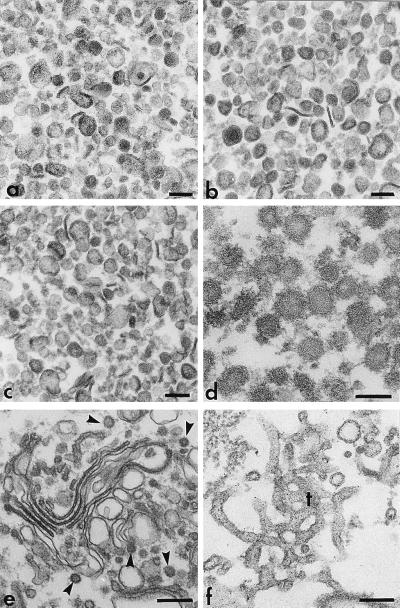
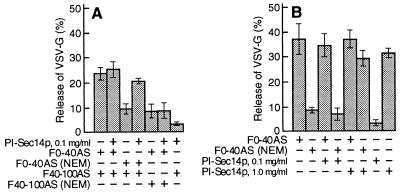
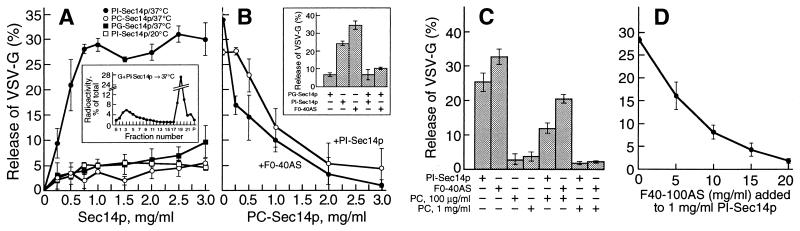
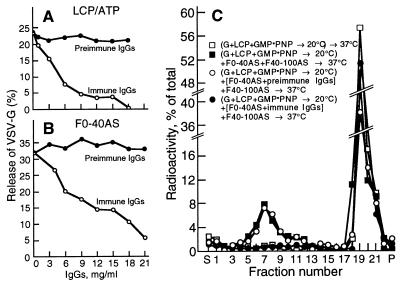
We identified the phosphatidylinositol transfer protein (PITP) as being responsible for a powerful latent, nucleotide-independent, Golgi-vesiculating activity that is present in the cytosol but is only manifested as an uncontrolled activity in a cytosolic protein subfraction, in which it is separated from regulatory components that appear to normally limit its action to the scission of COPI-coated buds from trans-Golgi network membranes. A specific anti-PITP antibody that recognizes the two mammalian PITP isoforms fully inhibited the capacity of the cytosol to support normal vesicle generation as well as the uncontrolled vesiculating activity manifested by the cytosolic protein subfraction. The phosphatidylinositol- (PI) loaded form of the yeast PITP, Sec14p, but not the phosphatidylcholine- (PC) loaded form of the protein, was capable of substituting for the cytosolic subfraction in promoting the scission of coated buds from the trans-Golgi network. At higher concentration, however, Sec14p, when loaded with PI, but not with PC or phosphatidylglycerol, caused by itself an indiscriminate vesiculation of uncoated Golgi membranes that could be suppressed by PC-Sec14p, which also suppresses the uncontrolled vesiculation caused by the cytosolic subfraction. We propose that, by delivering PI to specific sites in the Golgi membrane near the necks of coated buds, PITP induces local changes in the organization of the lipid bilayer, possibly involving PI metabolites, that triggers the fusion of the ectoplasmic faces of the Golgi membrane necessary for the scission of COPI-coated vesicles.
Figures

Similar articles
-
In vitro generation from the trans-Golgi network of coatomer-coated vesicles containing sialylated vesicular stomatitis virus-G protein.Methods. 2000 Apr;20(4):437-54. doi: 10.1006/meth.2000.0957. Methods. 2000. PMID: 10720465
-
Coatomer, but not P200/myosin II, is required for the in vitro formation of trans-Golgi network-derived vesicles containing the envelope glycoprotein of vesicular stomatitis virus.Proc Natl Acad Sci U S A. 1998 Feb 3;95(3):1073-8. doi: 10.1073/pnas.95.3.1073. Proc Natl Acad Sci U S A. 1998. PMID: 9448287 Free PMC article.
-
A phosphatidylinositol 3-kinase and phosphatidylinositol transfer protein act synergistically in formation of constitutive transport vesicles from the trans-Golgi network.J Biol Chem. 1998 Apr 24;273(17):10349-54. doi: 10.1074/jbc.273.17.10349. J Biol Chem. 1998. PMID: 9553090
-
Mechanism of formation of post Golgi vesicles from TGN membranes: Arf-dependent coat assembly and PKC-regulated vesicle scission.Biocell. 1996 Dec;20(3):287-300. Biocell. 1996. PMID: 9031596 Review.
-
Mammalian phosphatidylinositol transfer proteins: emerging roles in signal transduction and vesicular traffic.Chem Phys Lipids. 1999 Apr;98(1-2):23-33. doi: 10.1016/s0009-3084(99)00015-8. Chem Phys Lipids. 1999. PMID: 10358925 Review.
Cited by
-
Phospholipid synthesis participates in the regulation of diacylglycerol required for membrane trafficking at the Golgi complex.J Biol Chem. 2011 Aug 12;286(32):28632-43. doi: 10.1074/jbc.M111.267534. Epub 2011 Jun 23. J Biol Chem. 2011. PMID: 21700701 Free PMC article.
-
Rapid isolation of extracellular vesicles from cell culture and biological fluids using a synthetic peptide with specific affinity for heat shock proteins.PLoS One. 2014 Oct 17;9(10):e110443. doi: 10.1371/journal.pone.0110443. eCollection 2014. PLoS One. 2014. PMID: 25329303 Free PMC article.
-
Amot recognizes a juxtanuclear endocytic recycling compartment via a novel lipid binding domain.J Biol Chem. 2010 Apr 16;285(16):12308-20. doi: 10.1074/jbc.M109.096230. Epub 2010 Jan 14. J Biol Chem. 2010. PMID: 20080965 Free PMC article.
-
Genetics coupled to quantitative intact proteomics links heritable aphid and endosymbiont protein expression to circulative polerovirus transmission.J Virol. 2011 Mar;85(5):2148-66. doi: 10.1128/JVI.01504-10. Epub 2010 Dec 15. J Virol. 2011. PMID: 21159868 Free PMC article.
-
Mutations in Cog7 affect Golgi structure, meiotic cytokinesis and sperm development during Drosophila spermatogenesis.J Cell Sci. 2012 Nov 15;125(Pt 22):5441-52. doi: 10.1242/jcs.108878. Epub 2012 Sep 3. J Cell Sci. 2012. PMID: 22946051 Free PMC article.
References
-
- Barlowe C, Orci L, Yeung T, Hosobuchi M, Hamamoto S, Salama N, Rexach M F, Ravazzola M, Amherdt M, Schekman R. Cell. 1994;77:895–907. - PubMed
-
- Serafini T, Stenbeck G, Brecht A, Lottspeich F, Orci L, Rothman J E, Wieland F T. Nature (London) 1991;349:215–220. - PubMed
-
- Donaldson J G, Finazzi D, Klausner R D. Nature (London) 1992;360:350–352. - PubMed
-
- Helms J B, Rothman J E. Nature (London) 1992;360:352–354. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
