

Mad2 transiently associates with an APC/p55Cdc complex during mitosis
- PMID: 9736712
- PMCID: PMC21618
- DOI: 10.1073/pnas.95.19.11193
Mad2 transiently associates with an APC/p55Cdc complex during mitosis
Abstract
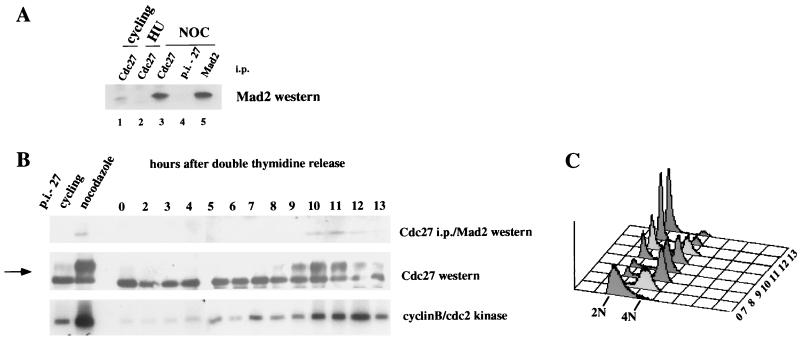
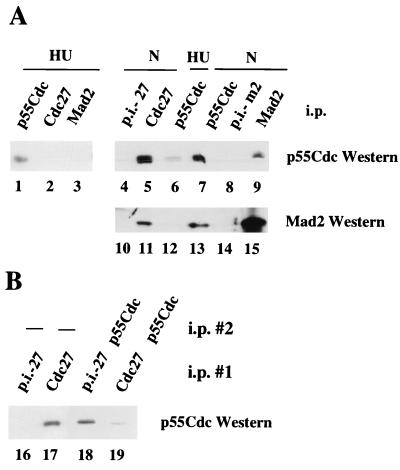
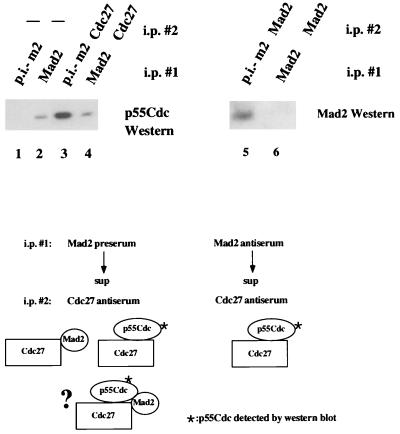
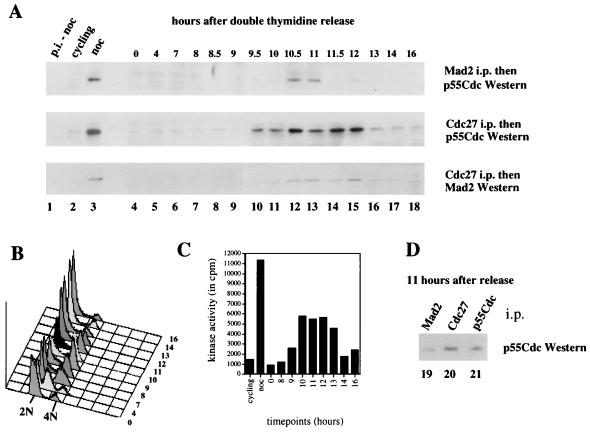
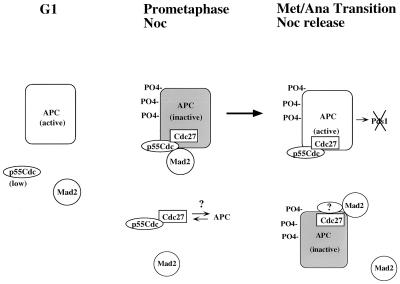
Activation of the mitotic checkpoint pathway in response to mitotic spindle damage in eukaryotic cells delays the exit from mitosis in an attempt to prevent chromosome missegregation. One component of this pathway, hsMad2, has been shown in mammalian cells to physically associate with components of a ubiquitin ligase activity (termed the anaphase promoting complex or APC) when the checkpoint is activated, thereby preventing the degradation of inhibitors of the mitotic exit machinery. In the present report, we demonstrate that the inhibitory association between Mad2 and the APC component Cdc27 also takes place transiently during the early stages of a normal mitosis and is lost before mitotic exit. We also show that Mad2 associates with the APC regulatory protein p55Cdc in mammalian cells as has been reported in yeast. In contrast, however, this complex is present only in nocodazole-arrested or early mitotic cells and is associated with the APC as a Mad2/p55Cdc/Cdc27 ternary complex. Evidence for a Mad2/Cdc27 complex that forms independent of p55Cdc also is presented. These results suggest a model for the regulation of the APC by Mad2 and may explain how the spindle assembly checkpoint apparatus controls the timing of mitosis under normal growth conditions.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
