

A novel DNA-binding protein bound to the mitochondrial inner membrane restores the null mutation of mitochondrial histone Abf2p in Saccharomyces cerevisiae
- PMID: 9742088
- PMCID: PMC109157
- DOI: 10.1128/MCB.18.10.5712
A novel DNA-binding protein bound to the mitochondrial inner membrane restores the null mutation of mitochondrial histone Abf2p in Saccharomyces cerevisiae
Abstract
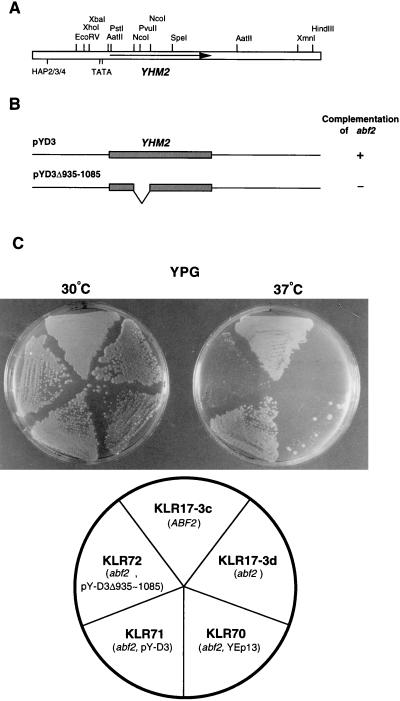
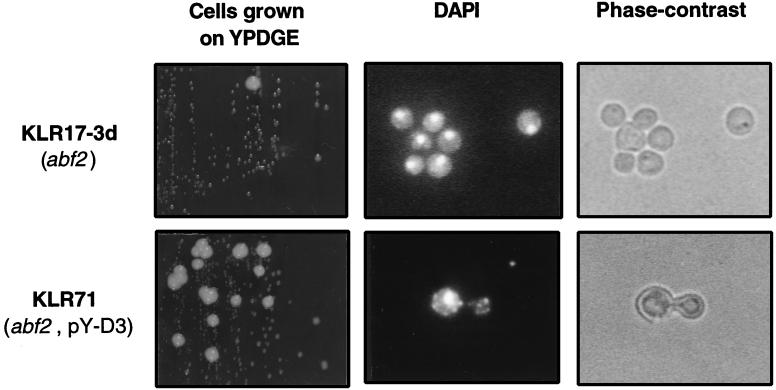
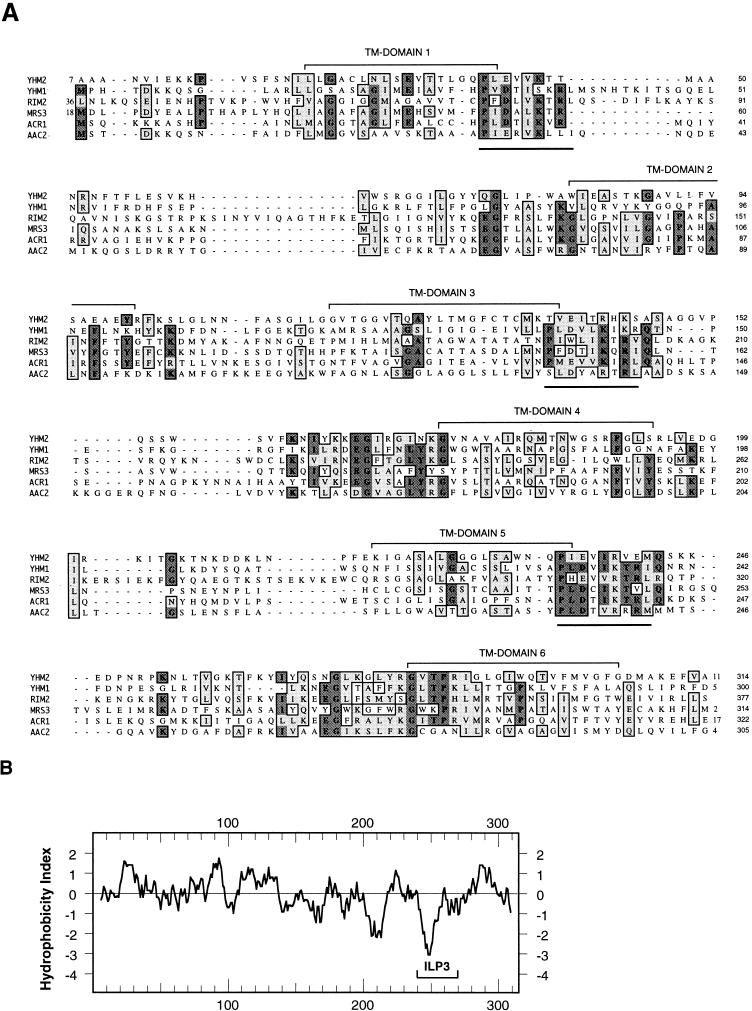
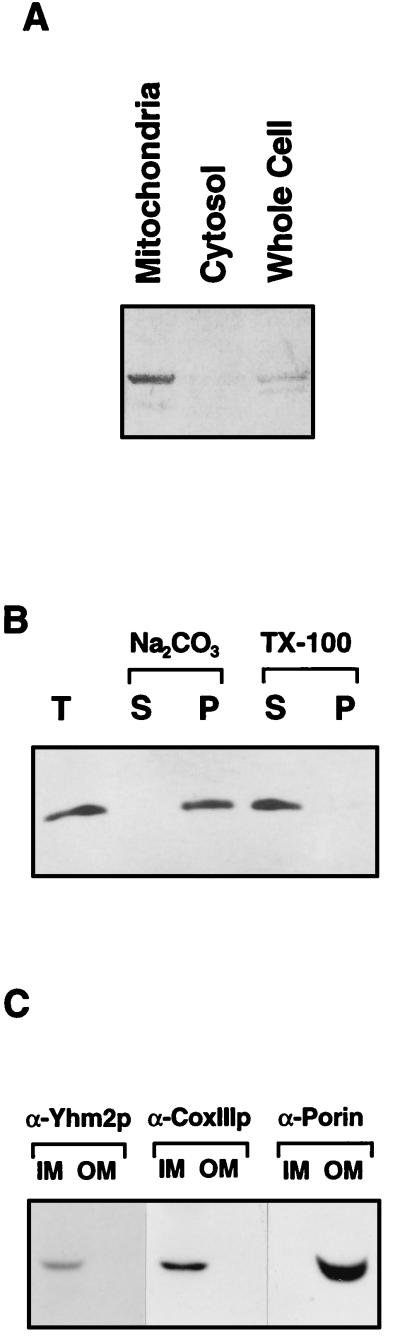
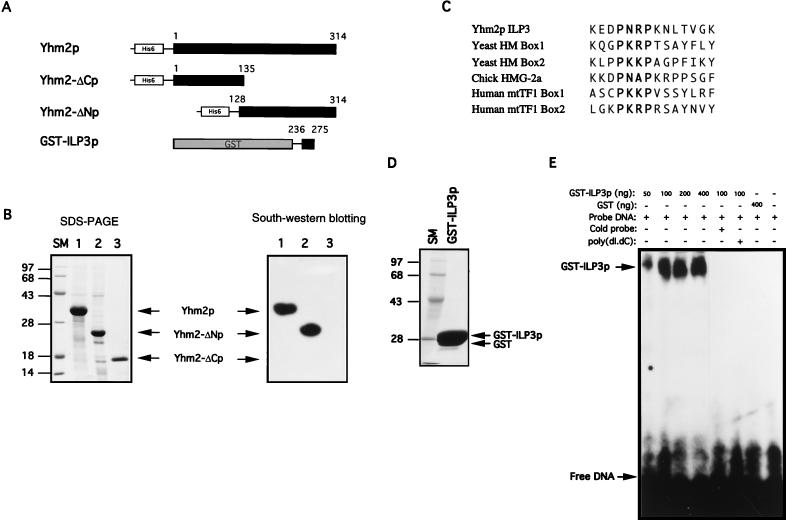
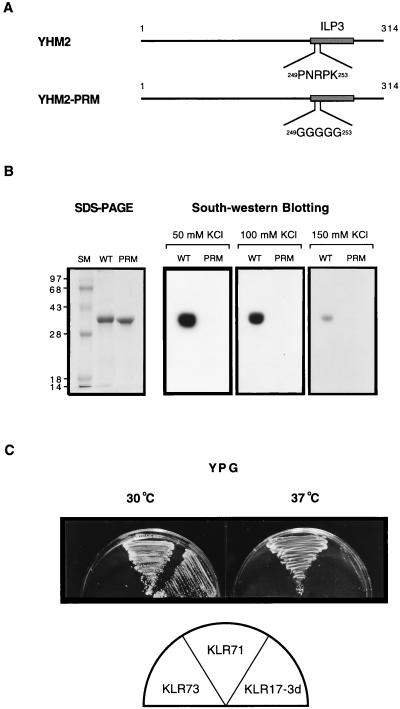
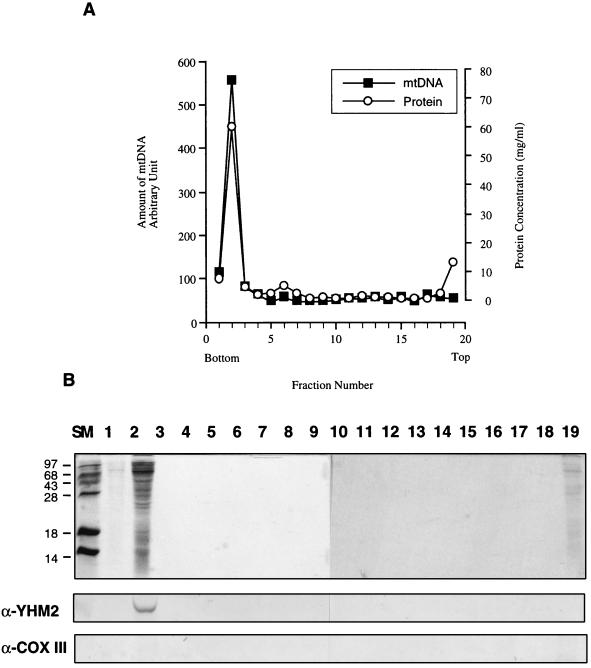
The yeast mitochondrial HMG-box protein, Abf2p, is essential for maintenance of the mitochondrial genome. To better understand the role of Abf2p in the maintenance of the mitochondrial chromosome, we have isolated a multicopy suppressor (YHM2) of the temperature-sensitive defect associated with an abf2 null mutation. The function of Yhm2p was characterized at the molecular level. Yhm2p has 314 amino acid residues, and the deduced amino acid sequence is similar to that of a family of mitochondrial carrier proteins. Yhm2p is localized in the mitochondrial inner membrane and is also associated with mitochondrial DNA in vivo. Yhm2p exhibits general DNA-binding activity in vitro. Thus, Yhm2p appears to be novel in that it is a membrane-bound DNA-binding protein. A sequence that is similar to the HMG DNA-binding domain is important for the DNA-binding activity of Yhm2p, and a mutation in this region abolishes the ability of YHM2 to suppress the temperature-sensitive defect of respiration of the abf2 null mutant. Disruption of YHM2 causes a significant growth defect in the presence of nonfermentable carbon sources such as glycerol and ethanol, and the cells have defects in respiration as determined by 2,3,5,-triphenyltetrazolium chloride staining. Yhm2p may function as a member of the protein machinery for the mitochondrial inner membrane attachment site of mitochondrial DNA during replication and segregation of mitochondrial genomes.
Figures
References
-
- Aquila H, Link T A, Klingenberg M. Solute carriers involved in energy transfer of mitochondria form a homologous protein family. FEBS Lett. 1987;212:1–9. - PubMed
-
- Cote C, Poirier J, Boulet D. Expression of the mammalian mitochondrial genome. Stability of mitochondrial translation products as a function of membrane potential. J Biol Chem. 1989;264:8487–8490. - PubMed
-
- Daum G, Bohni P C, Schatz G. Import of proteins into mitochondria. Energy-dependent uptake of precursors by isolated mitochondria. J Biol Chem. 1982;257:13028–13035. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases