

Macroscopic and microscopic properties of a cloned glutamate transporter/chloride channel
- PMID: 9742136
- PMCID: PMC6793006
- DOI: 10.1523/JNEUROSCI.18-19-07650.1998
Macroscopic and microscopic properties of a cloned glutamate transporter/chloride channel
Abstract
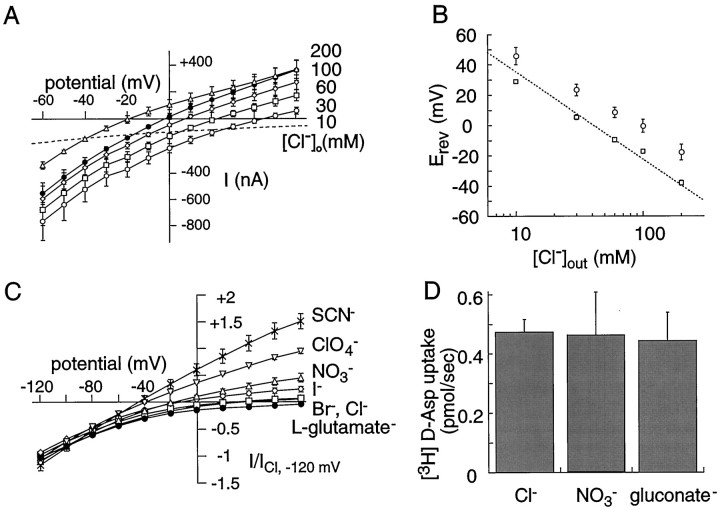
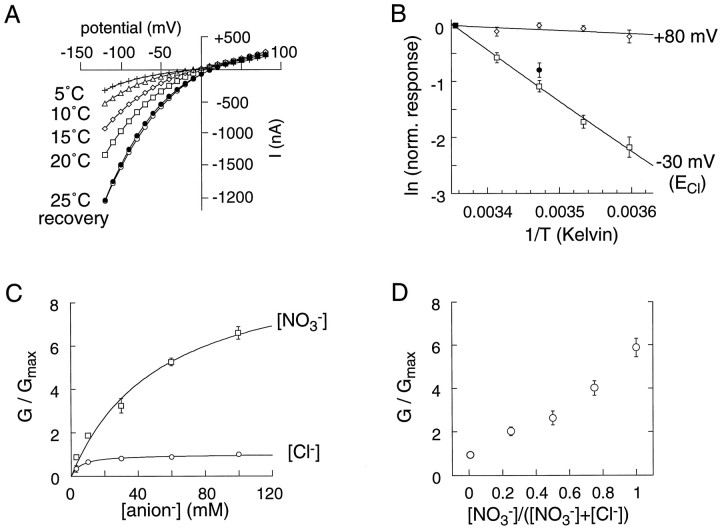
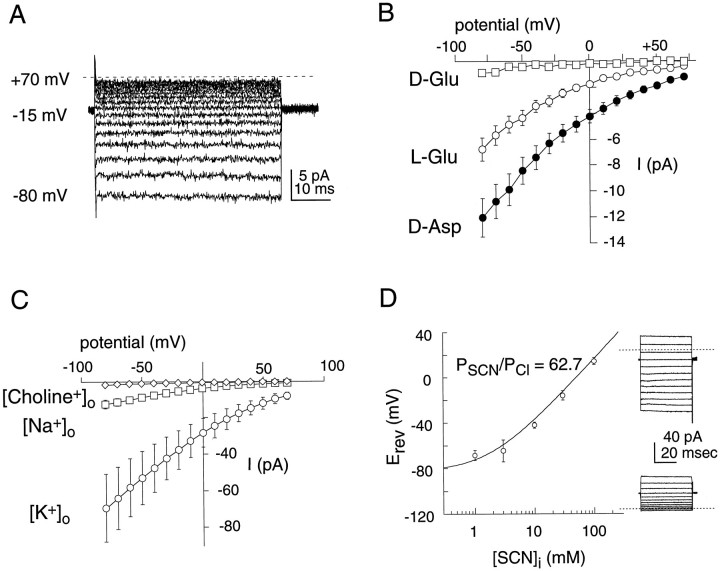
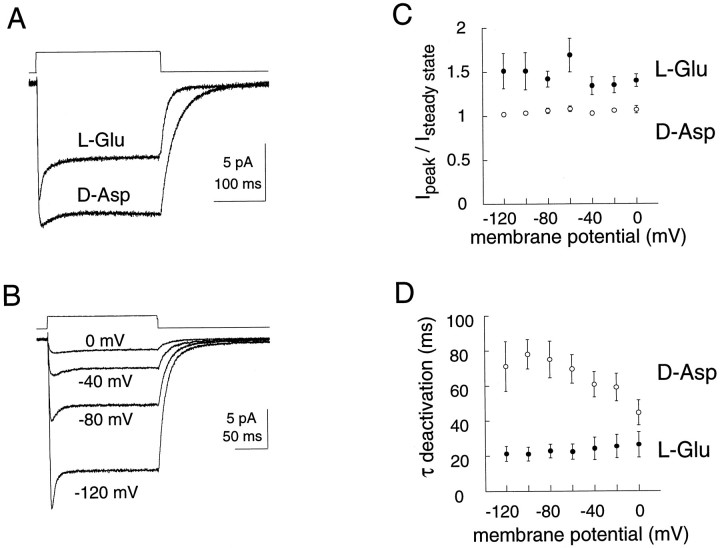
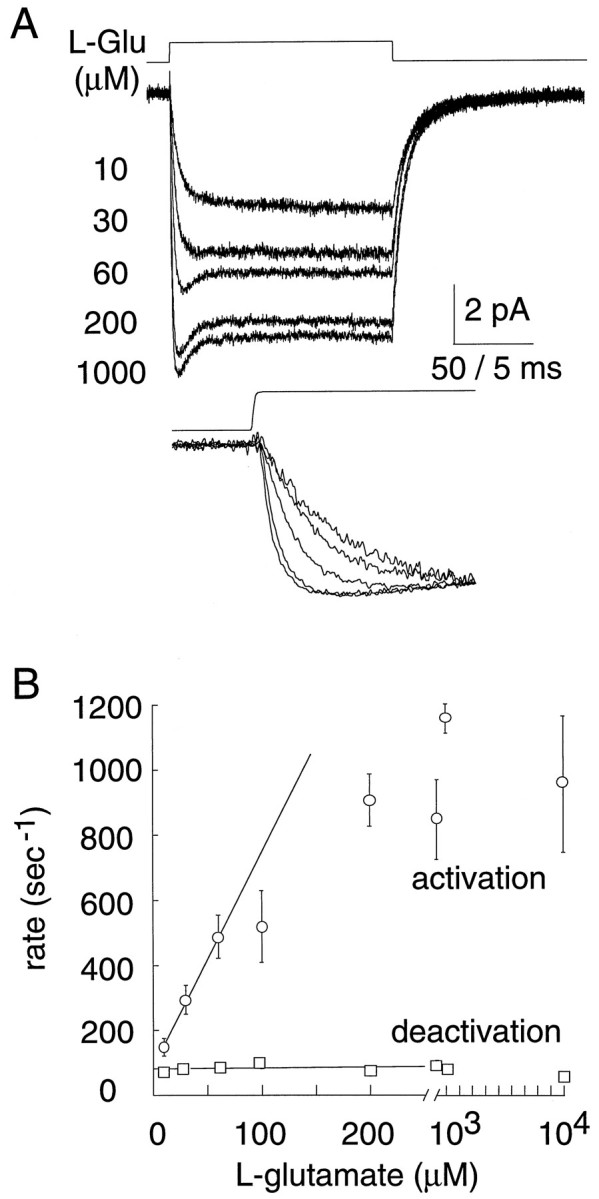
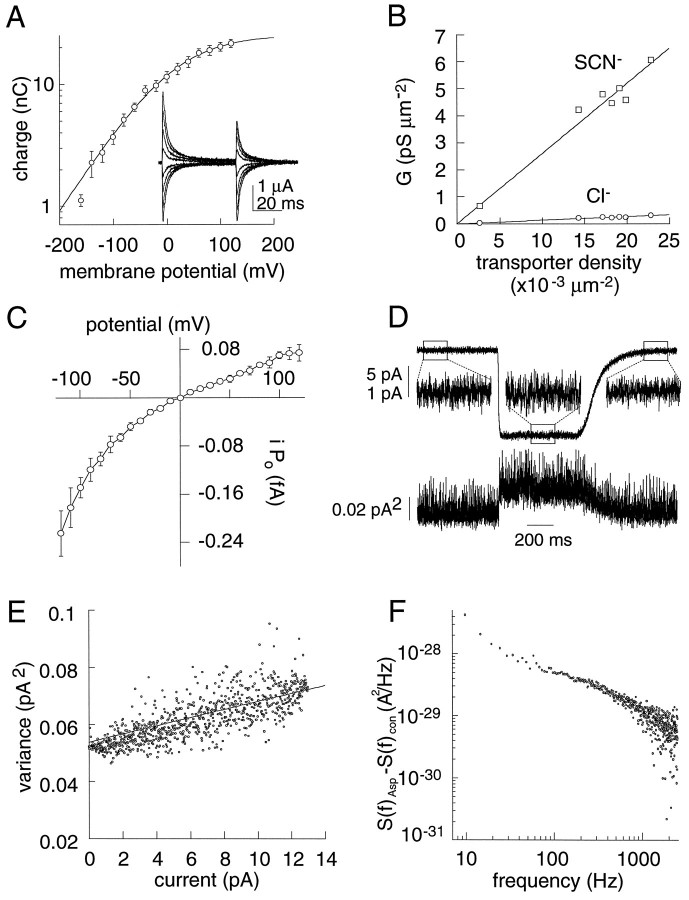
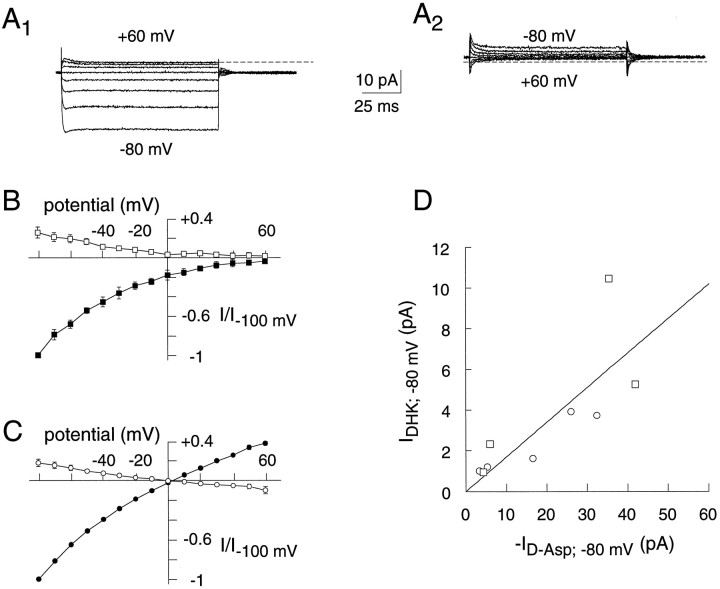
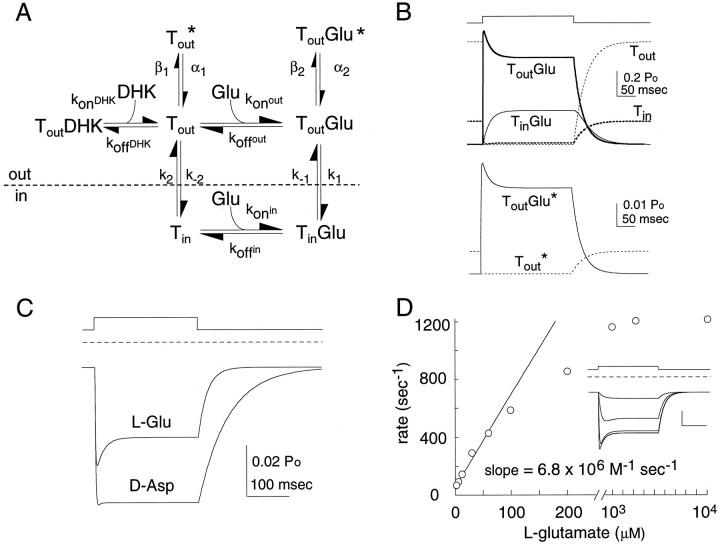
The behavior of a Cl- channel associated with a glutamate transporter was studied using intracellular and patch recording techniques in Xenopus oocytes injected with human EAAT1 cRNA. Channels could be activated by application of glutamate to either face of excised membrane patches. The channel exhibited strong selectivity for amphipathic anions and had a minimum pore diameter of approximately 5A. Glutamate flux exhibited a much greater temperature dependence than Cl- flux. Stationary and nonstationary noise analysis was consistent with a sub-femtosiemen Cl- conductance and a maximum channel Po << 1. The glutamate binding rate was similar to estimates for receptor binding. After glutamate binding, channels activated rapidly followed by a relaxation phase. Differences in the macroscopic kinetics of channels activated by concentration jumps of L-glutamate or D-aspartate were correlated with differences in uptake kinetics, indicating a close correspondence of channel gating to state transitions in the transporter cycle.
Figures
References
-
- Arriza JL, Kavanaugh MP, Fairman WA, Wu YN, Murdoch GH, North RA, Amara SG. Cloning and expression of a human neutral amino acid transporter with structural similarity to the glutamate transporter gene family. J Biol Chem. 1993;268:15329–15332. - PubMed
-
- Asztely F, Erdemli G, Kullmann DM. Extrasynaptic glutamate spillover in the hippocampus: dependence on temperature and the role of active glutamate uptake. Neuron. 1997;18:281–293. - PubMed
-
- Barbour B, Brew H, Attwell D. Electrogenic glutamate uptake in glial cells is activated by intracellular potassium. Nature. 1988;335:433–435. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources