

The cellular and subcellular localization of huntingtin-associated protein 1 (HAP1): comparison with huntingtin in rat and human
- PMID: 9742138
- PMCID: PMC6793025
- DOI: 10.1523/JNEUROSCI.18-19-07674.1998
The cellular and subcellular localization of huntingtin-associated protein 1 (HAP1): comparison with huntingtin in rat and human
Abstract
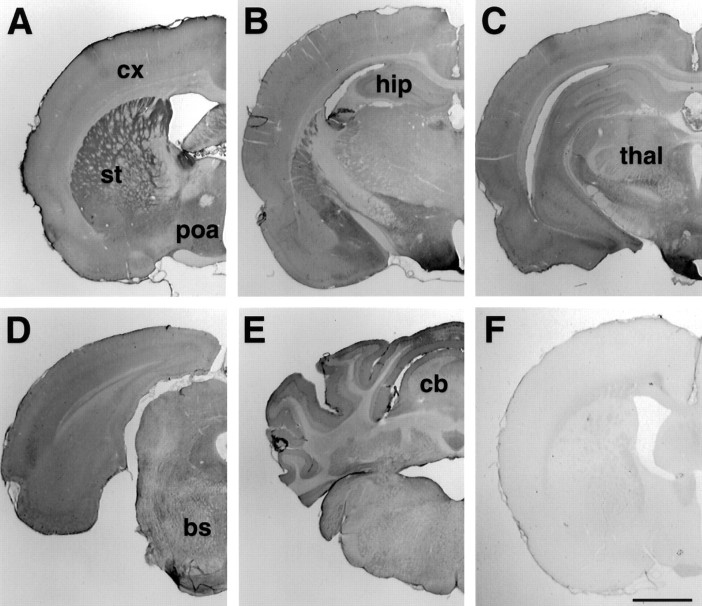
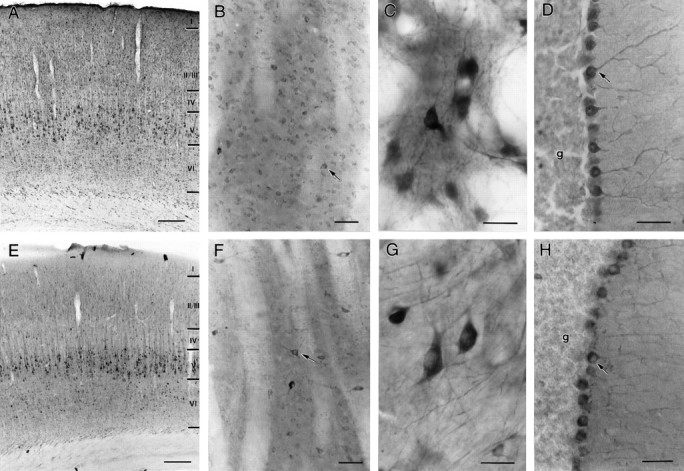
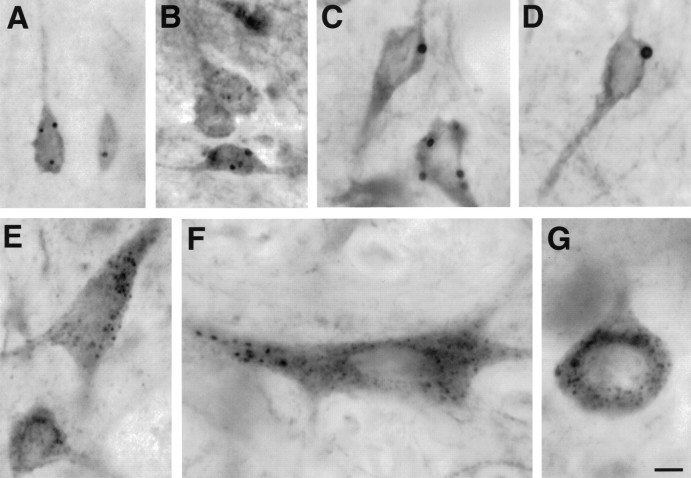
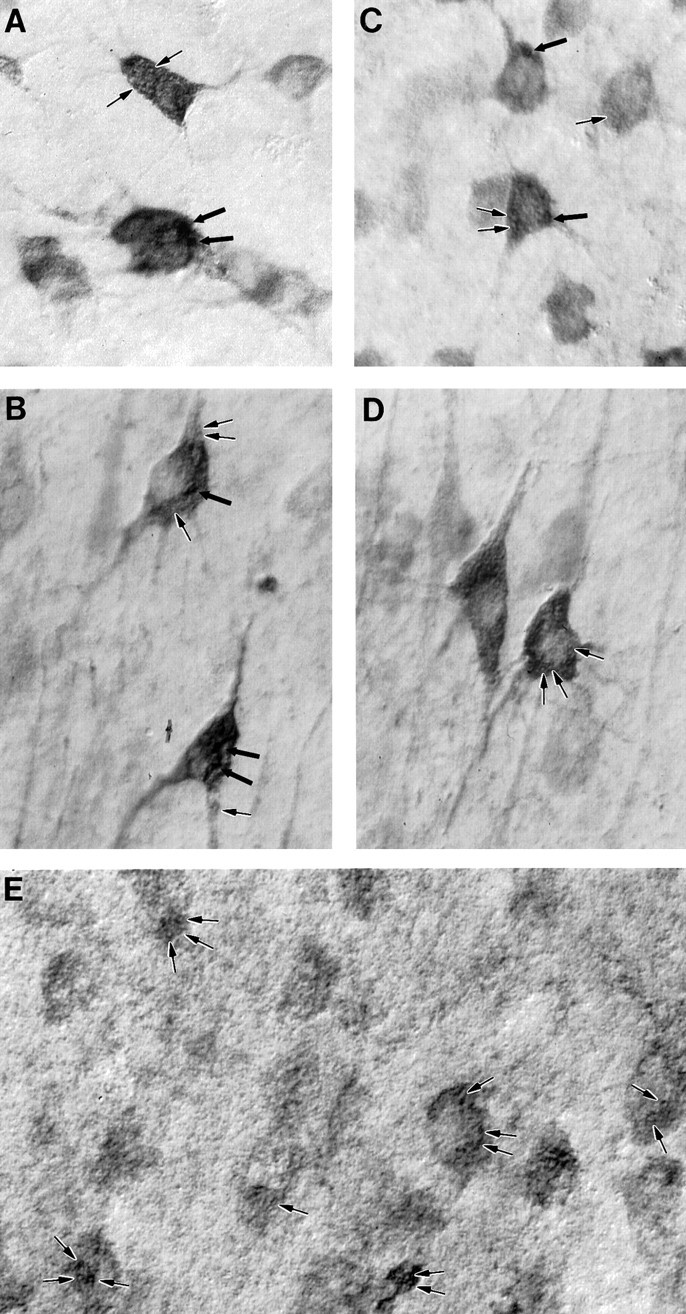
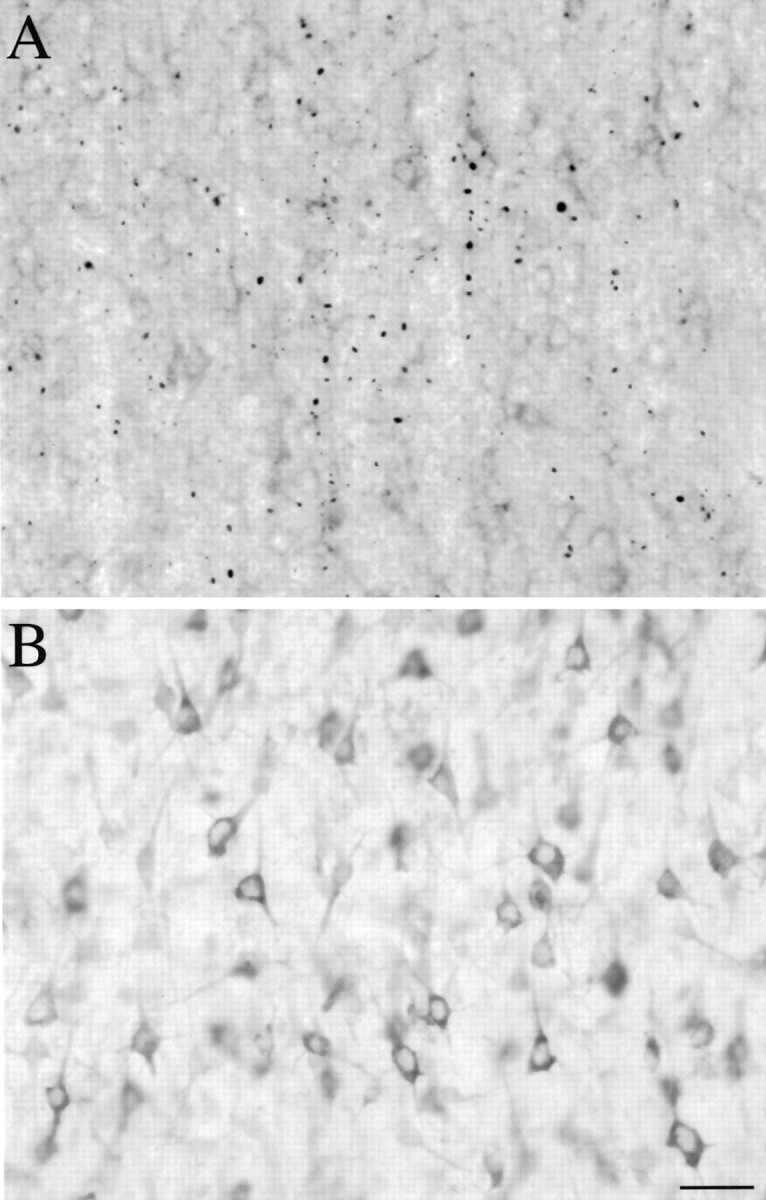
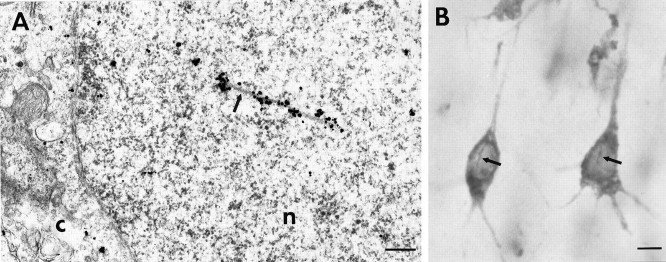
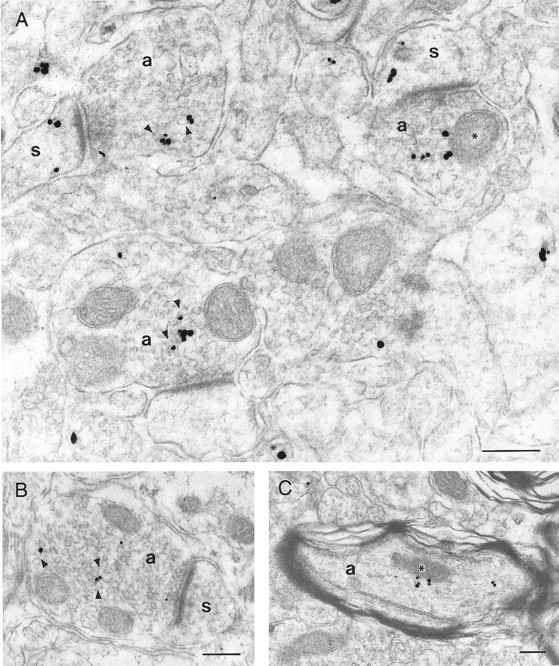
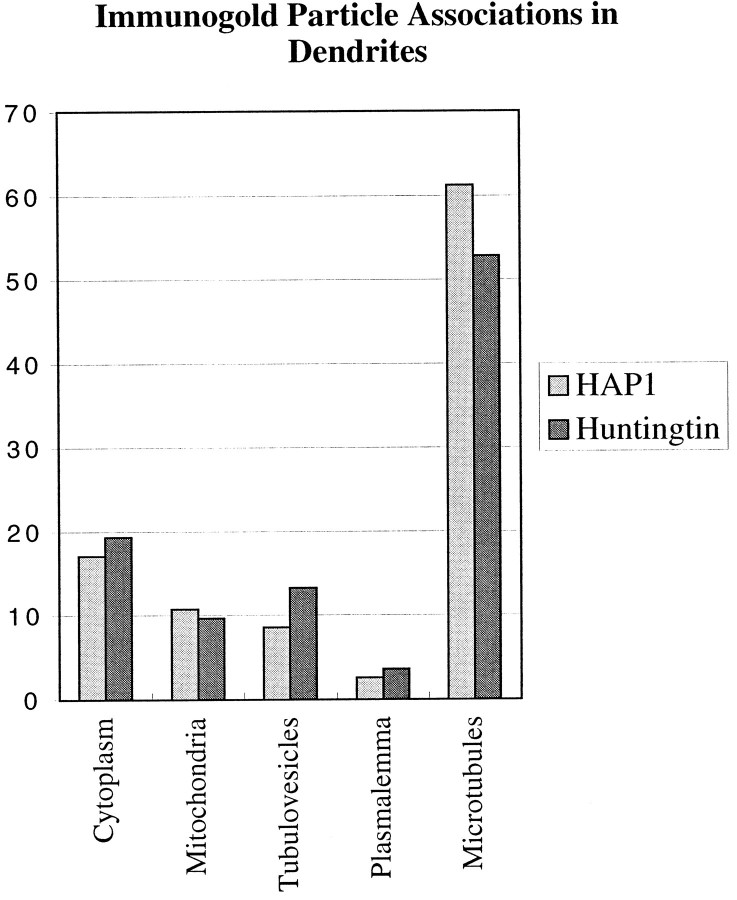
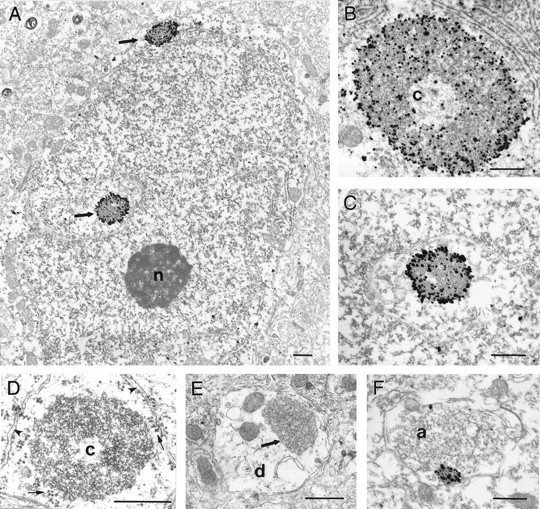
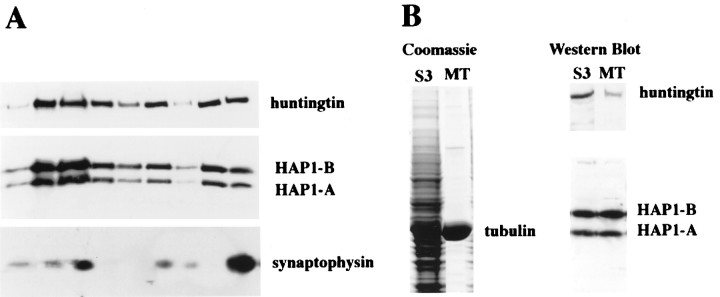
The cellular and subcellular distribution of HAP1 was examined in rat brain by light and electron microscopic immunocytochemistry and subcellular fractionation. HAP1 localization was also determined in human postmortem tissue from control and Huntington's disease (HD) cases by light microscopic immunocytochemistry. At the cellular level, the heterogeneity of HAP1 expression was similar to that of huntingtin; however, HAP1 immunoreactivity was more widespread. The subcellular distribution of HAP1 was examined using immunogold electron microscopy. Like huntingtin, HAP1 is a cytoplasmic protein that associates with microtubules and many types of membranous organelles, including mitochondria, endoplasmic reticulum, tubulovesicles, endosomal and lysosomal organelles, and synaptic vesicles. A quantitative comparison of the organelle associations of HAP1 and huntingtin showed them to be almost identical. Within HAP1-immunoreactive neurons in rat and human brain, populations of large and small immunoreactive puncta were visible by light microscopy. The large puncta, which were especially evident in the ventral forebrain, were intensely HAP1 immunoreactive. Electron microscopic analysis revealed them to be a type of nucleolus-like body, which has been named a stigmoid body, that may play a role in protein synthesis. The small puncta, less intensely labeled, were primarily mitochondria. These results indicate that the localization of HAP1 and huntingtin is more similar than previously appreciated and provide further evidence that HAP1 and huntingtin have localizations consistent with roles in intracellular transport. Our data also suggest, however, that HAP1 is not present in the abnormal intranuclear and neuritic aggregates containing the N-terminal fragment of mutant huntingtin that are found in HD brains.
Figures

References
-
- Becher MW, Kotzuk JA, Sharp AH, Davies SW, Bates GP, Price DL, Ross CA. Intranuclear neuronal inclusions in Huntington’s disease and dentatorubral and Pallidoluysian atrophy: correlation between the density of inclusions and IT15 CAG triplet repeat length. Neurobiol Dis. 1998;4:387–397. - PubMed
-
- Burke J, Enghild J, Martin M, Jou Y-S, Myers R, Roses A, Vance J, Strittmatter W. Huntingtin and DRPLA proteins selectively interact with the enzyme GAPDH. Nat Med. 1996;2:347–350. - PubMed
-
- Burry R, Vandre D, Hayes D. Silver enhancement of gold antibody probes in pre-embedding electron microscopic immunocytochemistry. J Histochem Cytochem. 1992;40:1849–1856. - PubMed
-
- Davies SW, Turmaine M, Cozens BA, DiFiglia M, Sharp AH, Ross CA, Scherzinger E, Wanker EE, Mangiarini L, Bates GP. Formation of neuronal intranuclear inclusions underlies the neurological dysfunction in mice transgenic for the HD mutation. Cell. 1997;90:537–548. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous