

Repression of yeast Ste12 transcription factor by direct binding of unphosphorylated Kss1 MAPK and its regulation by the Ste7 MEK
- PMID: 9744865
- PMCID: PMC317171
- DOI: 10.1101/gad.12.18.2887
Repression of yeast Ste12 transcription factor by direct binding of unphosphorylated Kss1 MAPK and its regulation by the Ste7 MEK
Abstract
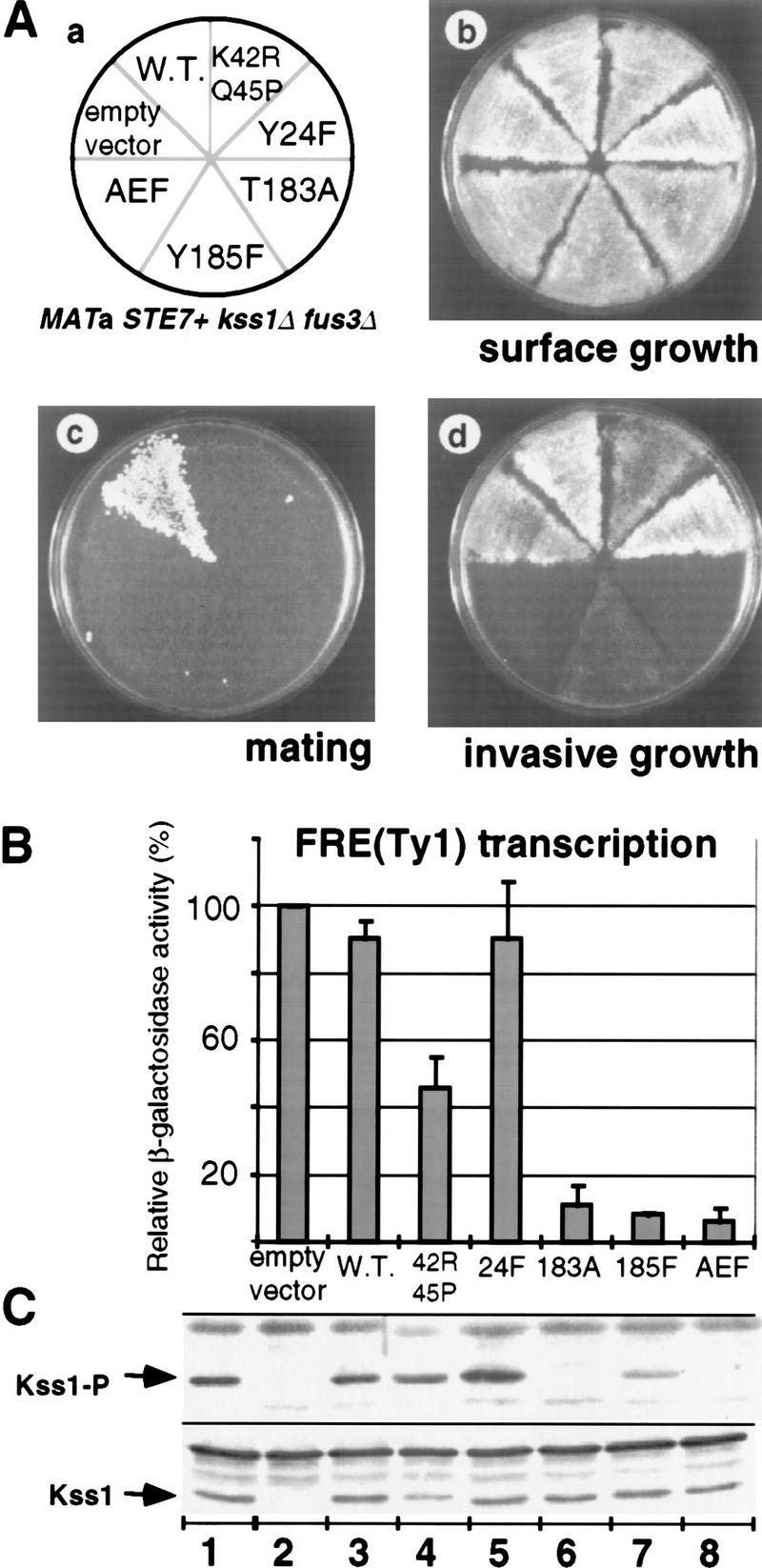
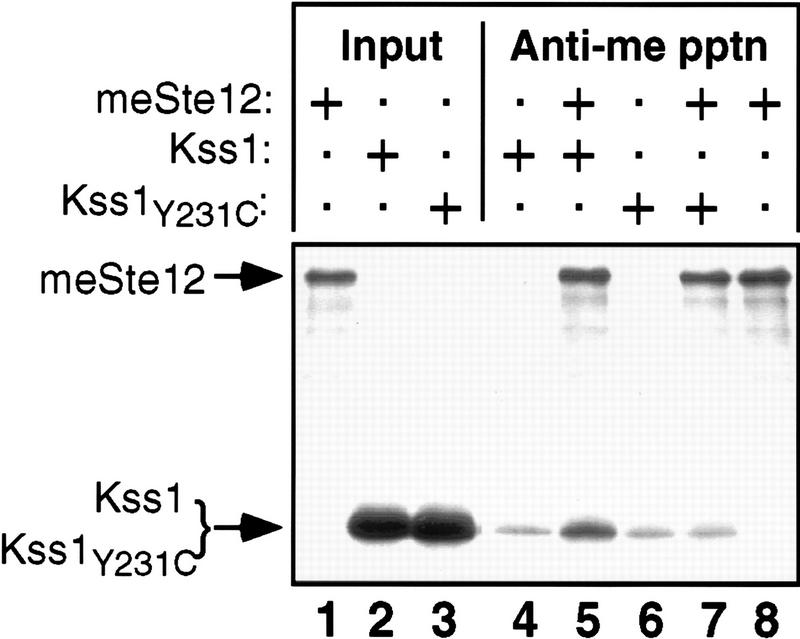
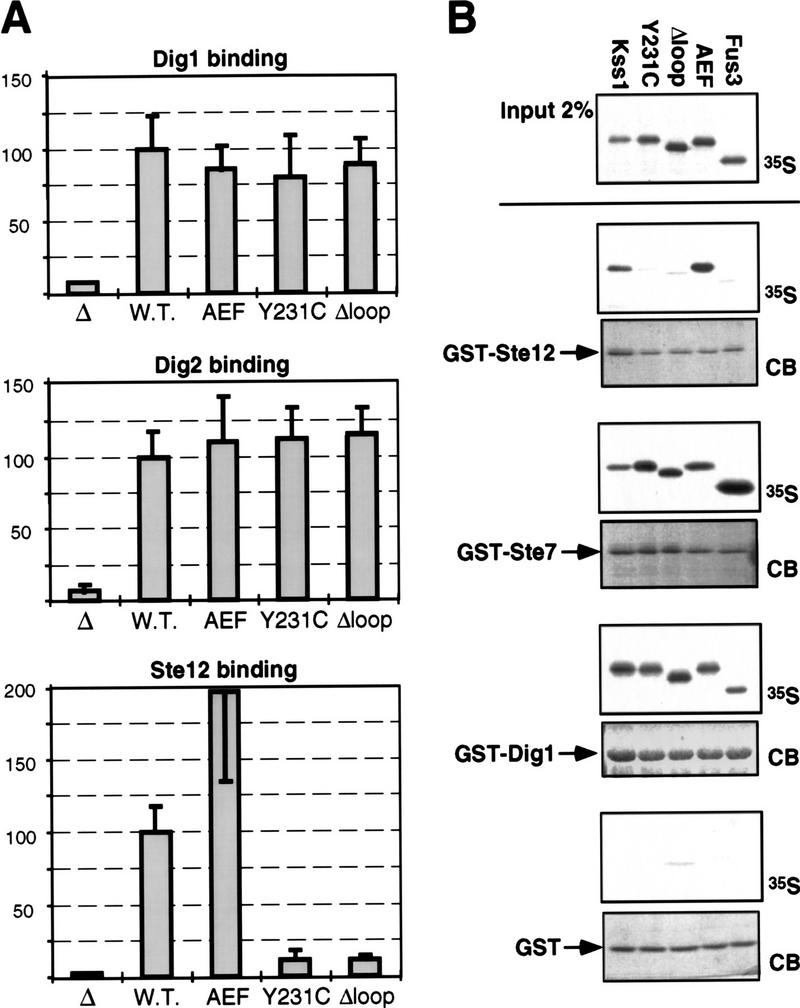
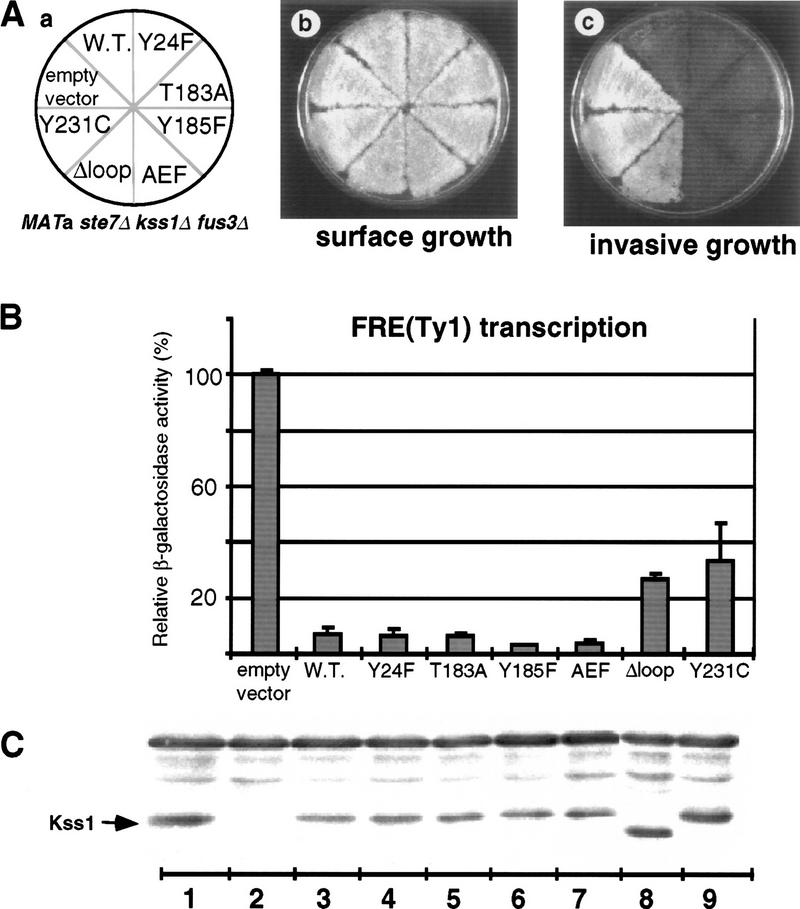
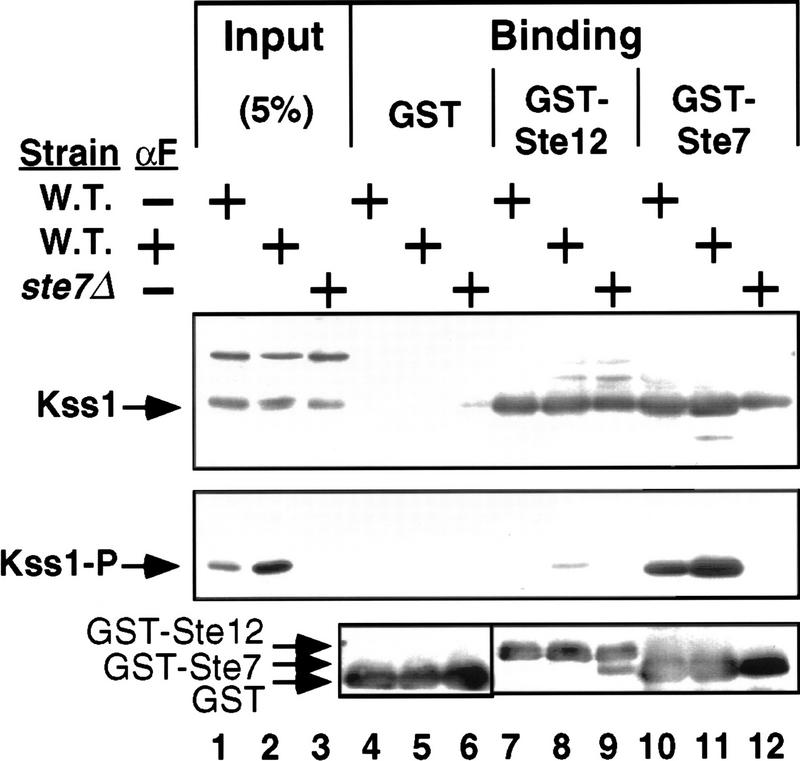
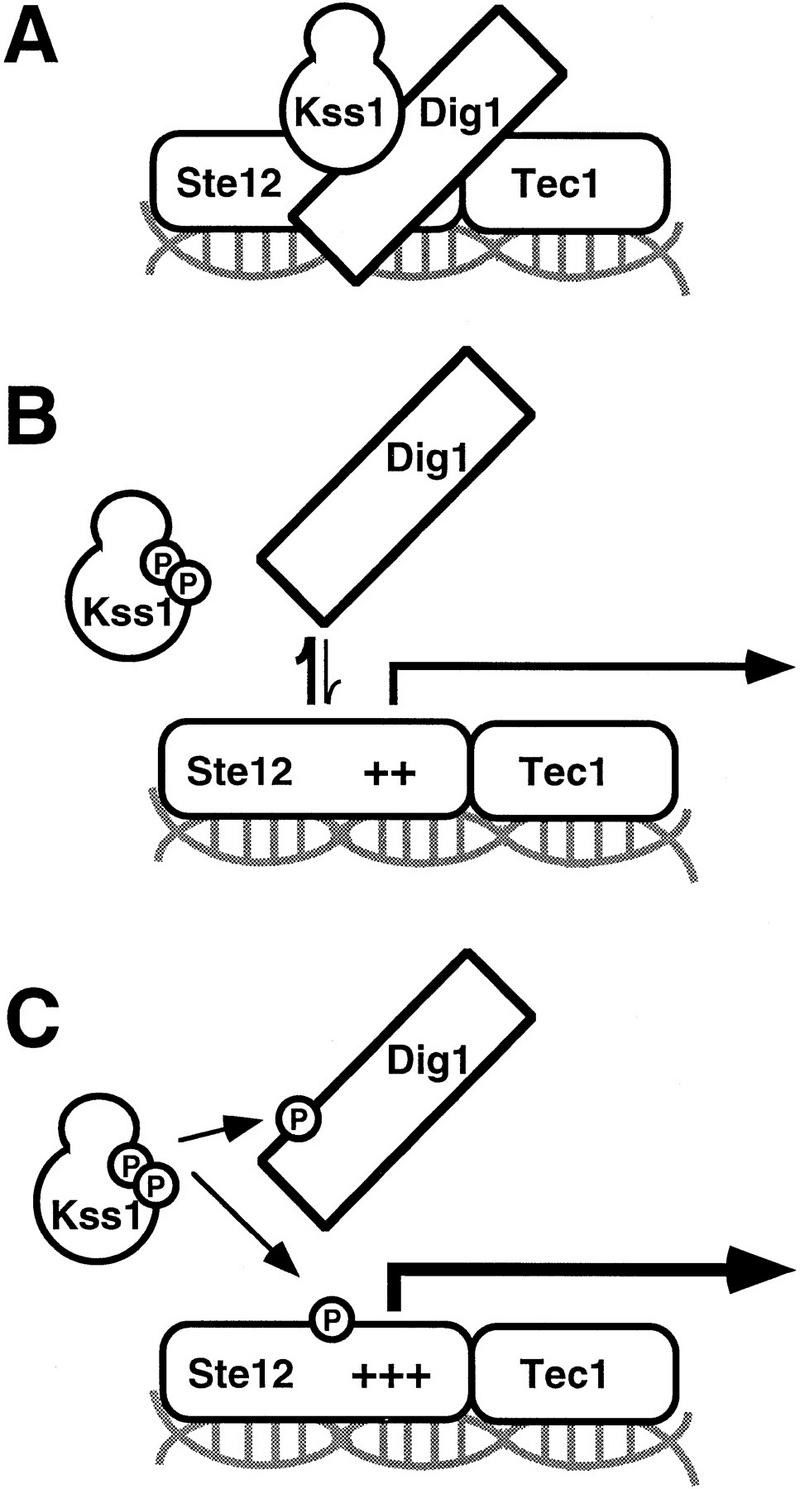
The mitogen-activated protein kinase (MAPK) Kss1 has a dual role in regulating filamentous (invasive) growth of the yeast Saccharomyces cerevisiae. The stimulatory function of Kss1 requires both its catalytic activity and its activation by the MAPK/ERK kinase (MEK) Ste7; in contrast, the inhibitory function of Kss1 requires neither. This study examines the mechanism by which Kss1 inhibits invasive growth, and how Ste7 action overcomes this inhibition. We found that unphosphorylated Kss1 binds directly to the transcription factor Ste12, that this binding is necessary for Kss1-mediated repression of Ste12, and that Ste7-mediated phosphorylation of Kss1 weakens Kss1-Ste12 interaction and relieves Kss1-mediated repression. Relative to Kss1, the MAPK Fus3 binds less strongly to Ste12 and is correspondingly a weaker inhibitor of invasive growth. Analysis of Kss1 mutants indicated that the activation loop of Kss1 controls binding to Ste12. Potent repression of a transcription factor by its physical interaction with the unactivated isoform of a protein kinase, and relief of this repression by activation of the kinase, is a novel mechanism for signal-dependent regulation of gene expression.
Figures

References
-
- Bardwell L, Thorner J. A conserved motif at the amino termini of MEKs might mediate high-affinity interaction with the cognate MAPKs. Trends Biochem Sci. 1996;1:373–374. - PubMed
-
- Bardwell L, Cook JG, Inouye CJ, Thorner J. Signal propagation and regulation in the mating pheromone response pathway of the yeast Saccharomyces cerevisiae. Devel Biol. 1994b;166:363–379. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous