

mei-W68 in Drosophila melanogaster encodes a Spo11 homolog: evidence that the mechanism for initiating meiotic recombination is conserved
- PMID: 9744869
- PMCID: PMC317166
- DOI: 10.1101/gad.12.18.2932
mei-W68 in Drosophila melanogaster encodes a Spo11 homolog: evidence that the mechanism for initiating meiotic recombination is conserved
Abstract
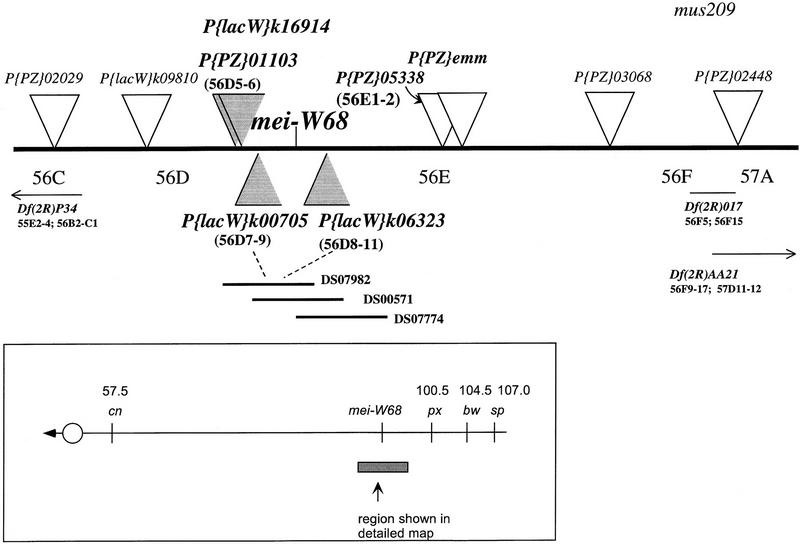
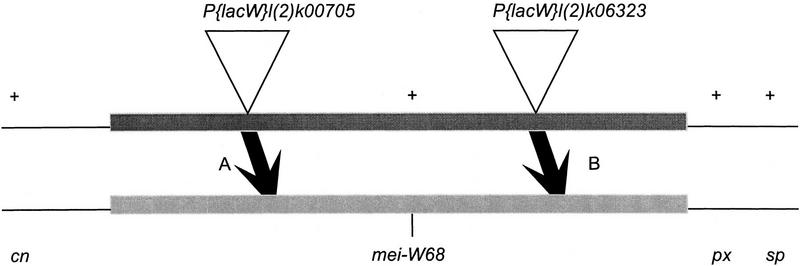
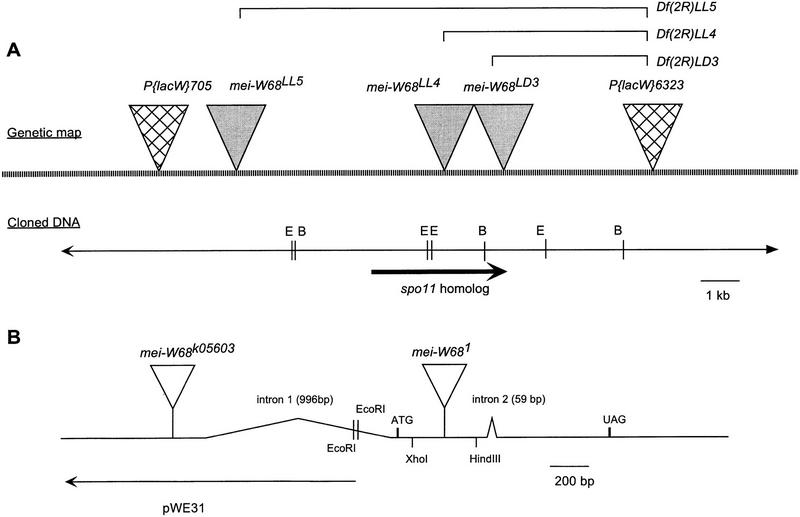
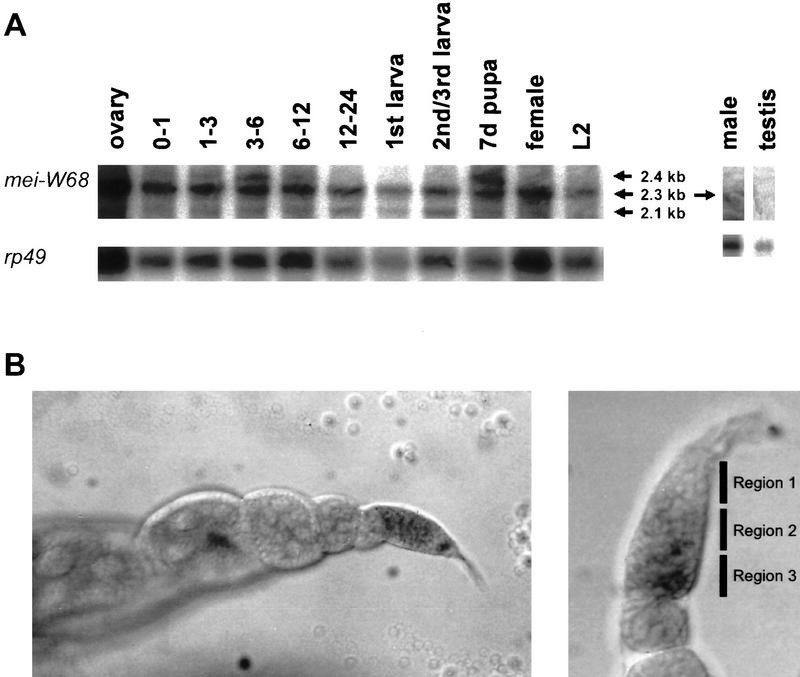
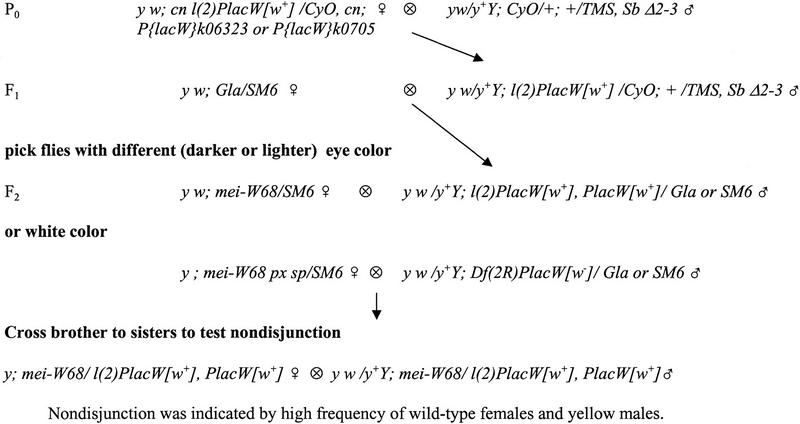
Meiotic recombination requires the action of several gene products in both Saccharomyces cerevisiae and Drosophila melanogaster. Genetic studies in D. melanogaster have shown that the mei-W68 gene is required for all meiotic gene conversion and crossing-over. We cloned mei-W68 using a new genetic mapping method in which P elements are used to promote crossing-over at their insertion sites. This resulted in the high-resolution mapping of mei-W68 to a <18-kb region that contains a homolog of the S. cerevisiae spo11 gene. Molecular analysis of several mutants confirmed that mei-W68 encodes an spo11 homolog. Spo11 and MEI-W68 are members of a family of proteins similar to a novel type II topoisomerase. On the basis of this and other lines of evidence, Spo11 has been proposed to be the enzymatic activity that creates the double-strand breaks needed to initiate meiotic recombination. This raises the possibility that recombination in Drosophila is also initiated by double-strand breaks. Although these homologous genes are required absolutely for recombination in both species, their roles differ in other respects. In contrast to spo11, mei-W68 is not required for synaptonemal complex formation and does have a mitotic role.
Figures

References
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases