

Activation of the MKK/ERK pathway during somatic cell mitosis: direct interactions of active ERK with kinetochores and regulation of the mitotic 3F3/2 phosphoantigen
- PMID: 9744882
- PMCID: PMC2141760
- DOI: 10.1083/jcb.142.6.1533
Activation of the MKK/ERK pathway during somatic cell mitosis: direct interactions of active ERK with kinetochores and regulation of the mitotic 3F3/2 phosphoantigen
Abstract
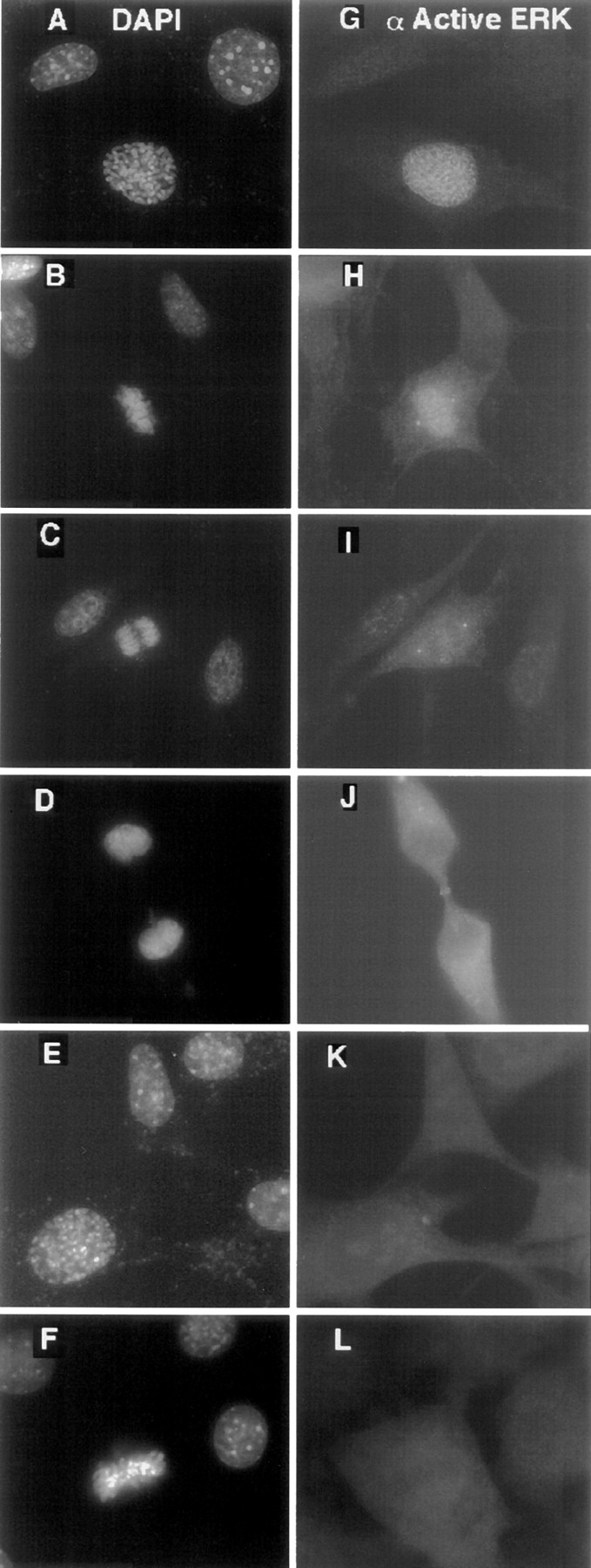
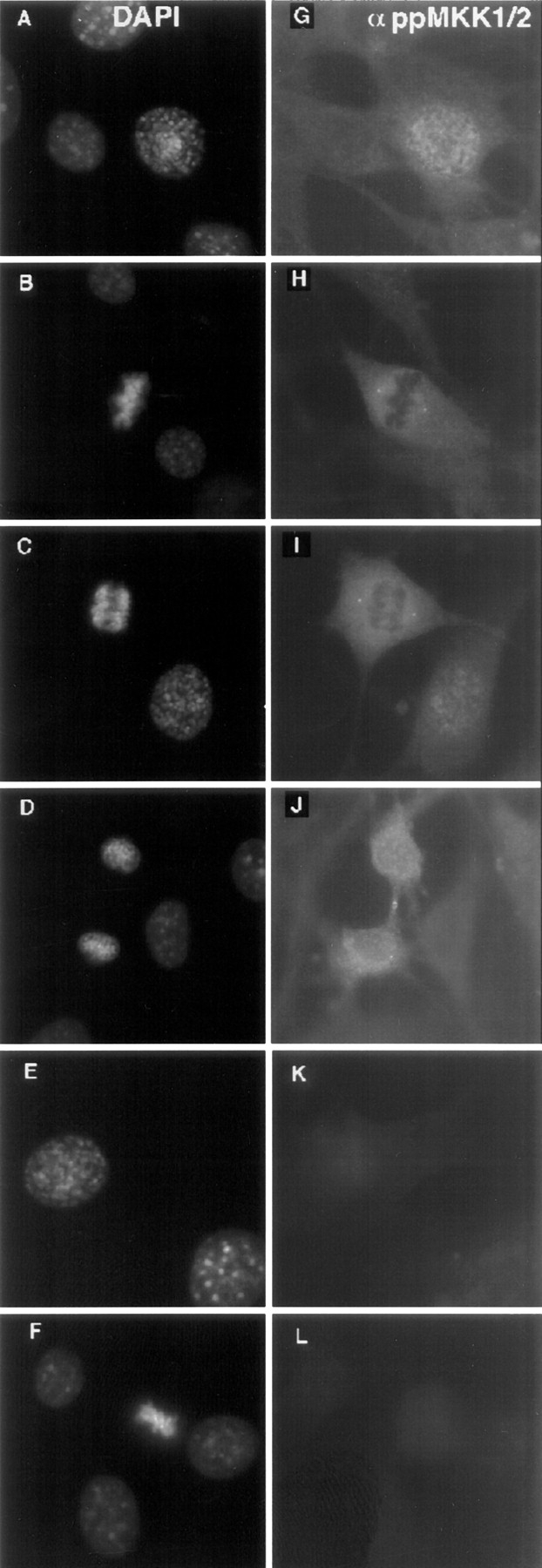
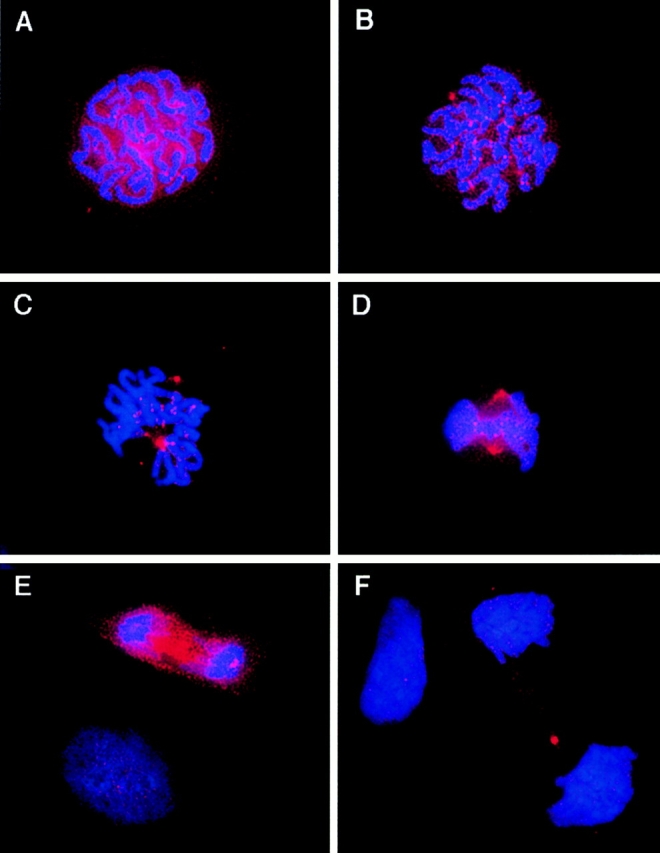
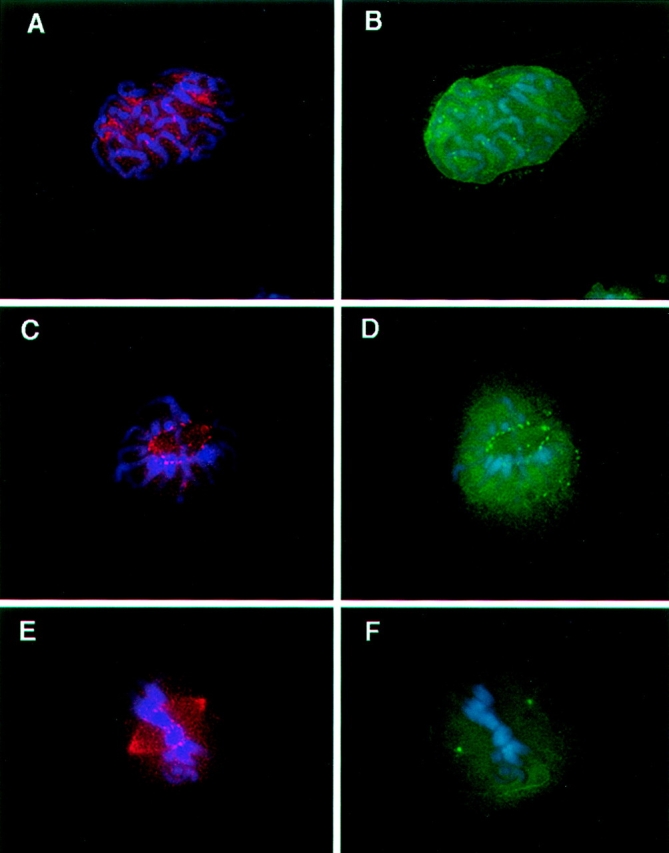
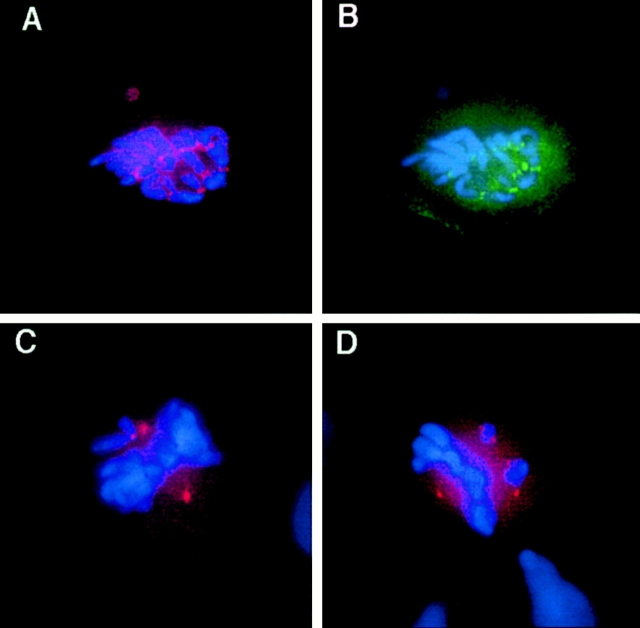
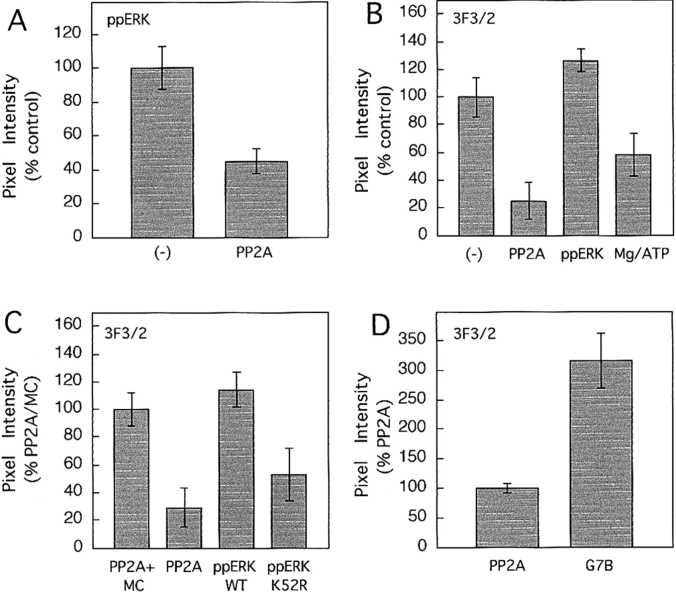
The mitogen-activated protein (MAP) kinase pathway, which includes extracellular signal-regulated protein kinases 1 and 2 (ERK1, ERK2) and MAP kinase kinases 1 and 2 (MKK1, MKK2), is well-known to be required for cell cycle progression from G1 to S phase, but its role in somatic cell mitosis has not been clearly established. We have examined the regulation of ERK and MKK in mammalian cells during mitosis using antibodies selective for active phosphorylated forms of these enzymes. In NIH 3T3 cells, both ERK and MKK are activated within the nucleus during early prophase; they localize to spindle poles between prophase and anaphase, and to the midbody during cytokinesis. During metaphase, active ERK is localized in the chromosome periphery, in contrast to active MKK, which shows clear chromosome exclusion. Prophase activation and spindle pole localization of active ERK and MKK are also observed in PtK1 cells. Discrete localization of active ERK at kinetochores is apparent by early prophase and during prometaphase with decreased staining on chromosomes aligned at the metaphase plate. The kinetochores of chromosomes displaced from the metaphase plate, or in microtubule-disrupted cells, still react strongly with the active ERK antibody. This pattern resembles that reported for the 3F3/2 monoclonal antibody, which recognizes a phosphoepitope that disappears with kinetochore attachment to the spindles, and has been implicated in the mitotic checkpoint for anaphase onset (Gorbsky and Ricketts, 1993. J. Cell Biol. 122:1311-1321). The 3F3/2 reactivity of kinetochores on isolated chromosomes decreases after dephosphorylation with protein phosphatase, and then increases after subsequent phosphorylation by purified active ERK or active MKK. These results suggest that the MAP kinase pathway has multiple functions during mitosis, helping to promote mitotic entry as well as targeting proteins that mediate mitotic progression in response to kinetochore attachment.
Figures







References
-
- Abrieu A, Lorca T, Labbe JC, Morin N, Keyse S, Doree M. MAP kinase does not inactivate, but rather prevents the cyclin degradation pathway from being turned on in Xenopusegg extracts. J Cell Sci. 1996;109:239–246. - PubMed
-
- Belmont LD, Mitchison TJ. Identification of a protein that interacts with tubulin dimers and increases the catastrophe rate of microtubules. Cell. 1996;84:623–631. - PubMed
-
- Beno WR, Brady LM, Bissonnette M, Davis BH. Protein kinase C and mitogen-activated protein kinase are required for 1,25-dihydroxy vitamin D3-stimulated Egr induction. J Biol Chem. 1995;270:3642–3647. - PubMed
-
- Brattsand G, Marklund U, Nylander K, Roos G, Gullberg M. Cell-cycle-regulated phosphorylation of oncoprotein 18 on Ser18, Ser25, and Ser38. Eur J Biochem. 1994;220:359–368. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
