

doi: 10.1073/pnas.95.20.11555.
RNA folding causes secondary structure rearrangement
Affiliations
- PMID: 9751704
- PMCID: PMC21679
- DOI: 10.1073/pnas.95.20.11555
Item in Clipboard
RNA folding causes secondary structure rearrangement
Proc Natl Acad Sci U S A.
.
Abstract
The secondary structure of the P5abc subdomain (a 56-nt RNA) of the Tetrahymena thermophila group I intron ribozyme has been determined by NMR. Its base pairing in aqueous solution in the absence of magnesium ions is significantly different from the RNA in a crystal but is consistent with thermodynamic predictions. On addition of magnesium ions, the RNA folds into a tertiary structure with greatly changed base pairing consistent with the crystal structure: three Watson-Crick base pairs, three G.U base pairs, and an extra-stable tetraloop are lost. The common assumption that RNA folds by first forming secondary structure and then forming tertiary interactions from the unpaired bases is not always correct.
Figures
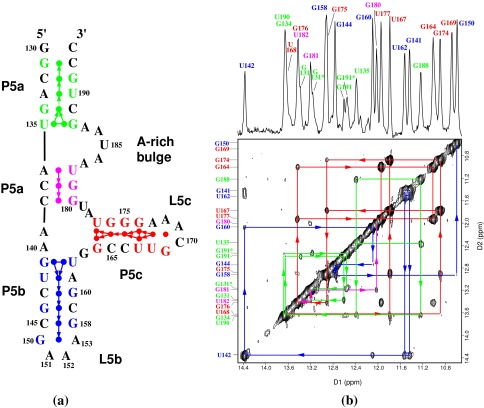
(a) The secondary structure of the
56-nt P5abc subdomain. The numbering is the same as in the P4–P6
domain. P5abc has 4 bp deleted from the P4–P6 domain between the
C145⋅G158 base pair of P5b and the L5b tetraloop GAAA (nucleotides
150–153); the 4-bp deletion causes the discontinuity in the numbering
of this region. The disk between each base pair represents the observed
imino proton of the base pair. The terminal G130 imino proton resonance
is not observed by NMR because of the fraying of the G130⋅C193
base pair. Each arrow represents an NOE connectivity between two imino
protons; the direction and color of the arrows match those of NOE walks
in the 2D spectrum shown in b. The dotted arrow between
the G134 and U190 imino protons indicates our inability to see the NOE
between them caused by the near identity of their chemical shifts.
(b) The 1D imino proton spectrum (Upper)
and 2D 120-ms NOESY spectrum (Lower) of 2.5 mM of P5abc
in 10 mM sodium phosphate and 0.01 mM EDTA (pH 6.4) in 90%
H2O/10% 2H2O at 10°C. The 1D
spectrum was acquired with 4K complex points and processed with a
25°-shifted sine bell squared window function. The 2D data were
acquired with 4K points in the D1 dimension and 512 points in the D2
dimension, and were processed with a 40°-shifted sine bell squared
function. The arrows connecting diagonal peaks and crosspeaks and the
NOE connectivities indicate the spatial arrangement of the imino
protons in the secondary structure. The NOE walks are color-coded to
the stems in the secondary structure. The 3′ inhomogeneity of the
molecule, which is caused by the presence of the N and N+1 species,
splits the chemical shifts of imino protons of G131, G191, and U190.
Peaks G131* and G191* are from the N+1 impurity molecule, a 57-mer. The
peak doubling stops at G134 and does not affect the rest of the
molecule.
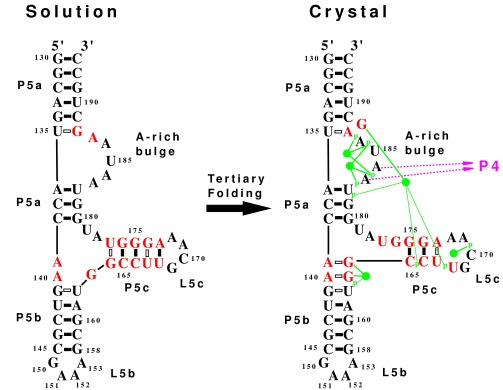
The secondary structures determined by NMR in
solution and by x-ray diffraction in a crystal are compared.
Nucleotides that change their base pairing are in red. The solution
structure represents the unfolded state, whereas the crystal structure
represents the folded state with tertiary interactions. Each solid bar
denotes a Watson–Crick base pair, and each hollow bar denotes a
non-Watson–Crick base pair. The tertiary interactions seen in the
crystal (10) include binding of A183 and A184 to the minor groove of P4
(shown as purple arrows). The green disks represent five magnesium ions
identified in the crystal (9, 10) that form direct hydrogen bonds
(thick green line) and water-mediated hydrogen bonds (thin green lines)
with phosphate oxygens (indicated by p) and guanine bases. The tertiary
folding causes significant secondary structure rearrangements,
including the loss of three G⋅U base pairs, the loss of a
GNRA tetraloop, the formation of a single base bulge, and the addition
of two G⋅A base pairs.
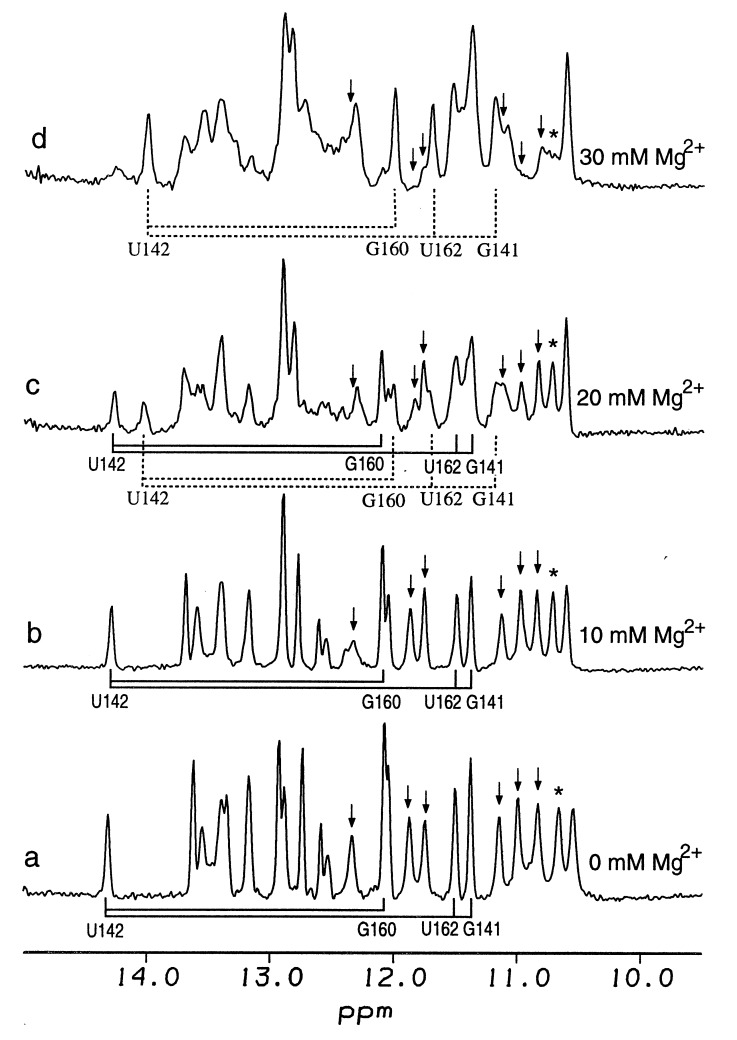
Mg2+-induced folding monitored by
imino proton NMR. Spectra of 0.4 mM of P5abc in 10 mM sodium phosphate
and 0.05 mM EDTA (pH 6.4) were measured at 20°C as a function of
added MgCl2. The total concentration of Mg2+
was 0 mM (a), 10 mM (b), 20 mM
(c), and 30 mM (d). Solid and dashed
lines represent the NOE connectivities as determined by 2D NOESY of 3
bp in stem P5b (G141⋅U162, U142⋅A161, and C143⋅G160) for
the unfolded and folded conformations, respectively. With the addition
of Mg2+, the four imino proton peaks of these base pairs
decrease, and a new set of four imino proton peaks with the same NOE
connectivities appear at different chemical shifts. This response is
characteristic of a slow exchange between the folded and unfolded
conformations. The percentages of folding estimated from the relative
intensity of U142 in the folded and unfolded states are ≈50% at 20
mM Mg2+ and 80% at 30 mM Mg2+. Six imino
proton peaks of three G⋅U pairs (G188⋅U135, G164⋅U177,
and G174⋅U167) disappear during the folding; the peaks are
indicated by arrows. NOESY spectra of c and
d show that these six bases no longer form G⋅U
pairs in the folded state. An asterisk indicates the sheared G⋅A
base pair of tetraloop L5c that disappears on folding.
Comment in
-
Native secondary structure formation in RNA may be a slave to tertiary folding.Proc Natl Acad Sci U S A. 1998 Sep 29;95(20):11506-8. doi: 10.1073/pnas.95.20.11506. Proc Natl Acad Sci U S A. 1998. PMID: 9751694 Free PMC article. Review. No abstract available.
References
-
- Gesteland R F, Atkins J F, editors. The RNA World. Plainview, NY: Cold Spring Harbor Lab. Press; 1993.
-
- Simons R W, Grunberg-Manago M, editors. RNA Structure and Function. Plainview, NY: Cold Spring Harbor Lab. Press; 1997.
-
- Jaeger J A, Zuker M, Turner D H. Biochemistry. 1990;29:10147–10158. - PubMed
-
- Celander D W, Cech T R. Science. 1991;251:401–407. - PubMed
-
- Banerjee A R, Jaeger J A, Turner D H. Biochemistry. 1993;32:153–163. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
