

The endoribonucleolytic N-terminal half of Escherichia coli RNase E is evolutionarily conserved in Synechocystis sp. and other bacteria but not the C-terminal half, which is sufficient for degradosome assembly
- PMID: 9751718
- PMCID: PMC21693
- DOI: 10.1073/pnas.95.20.11637
The endoribonucleolytic N-terminal half of Escherichia coli RNase E is evolutionarily conserved in Synechocystis sp. and other bacteria but not the C-terminal half, which is sufficient for degradosome assembly
Abstract
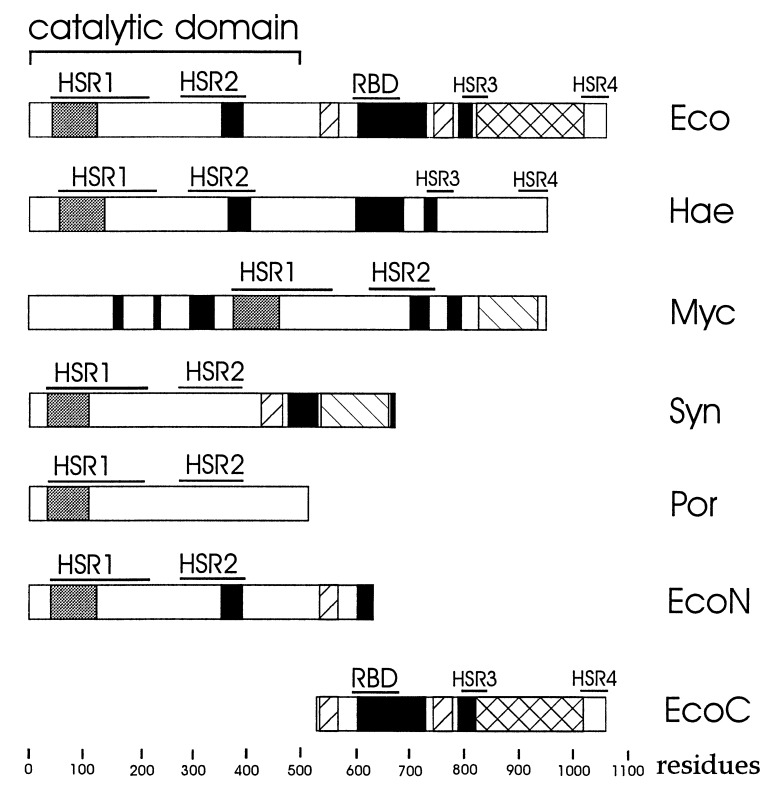
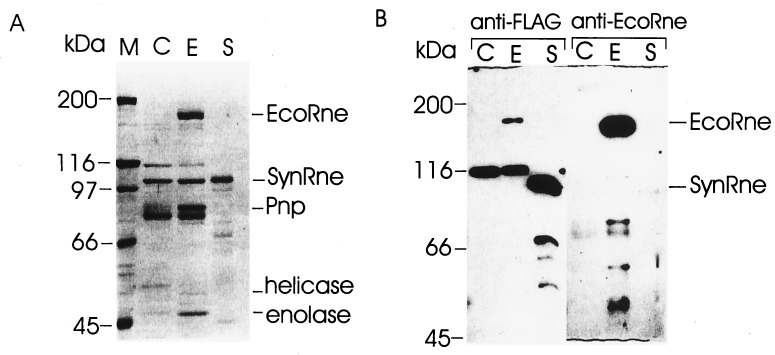
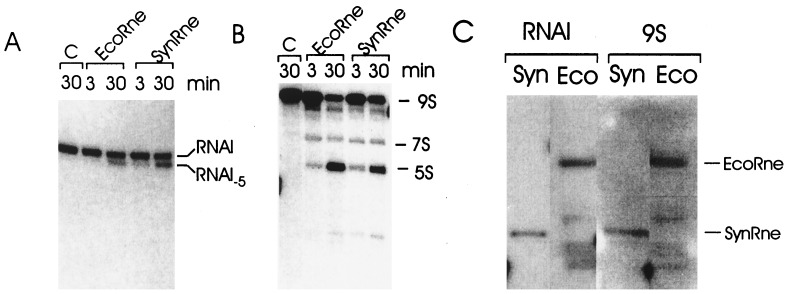
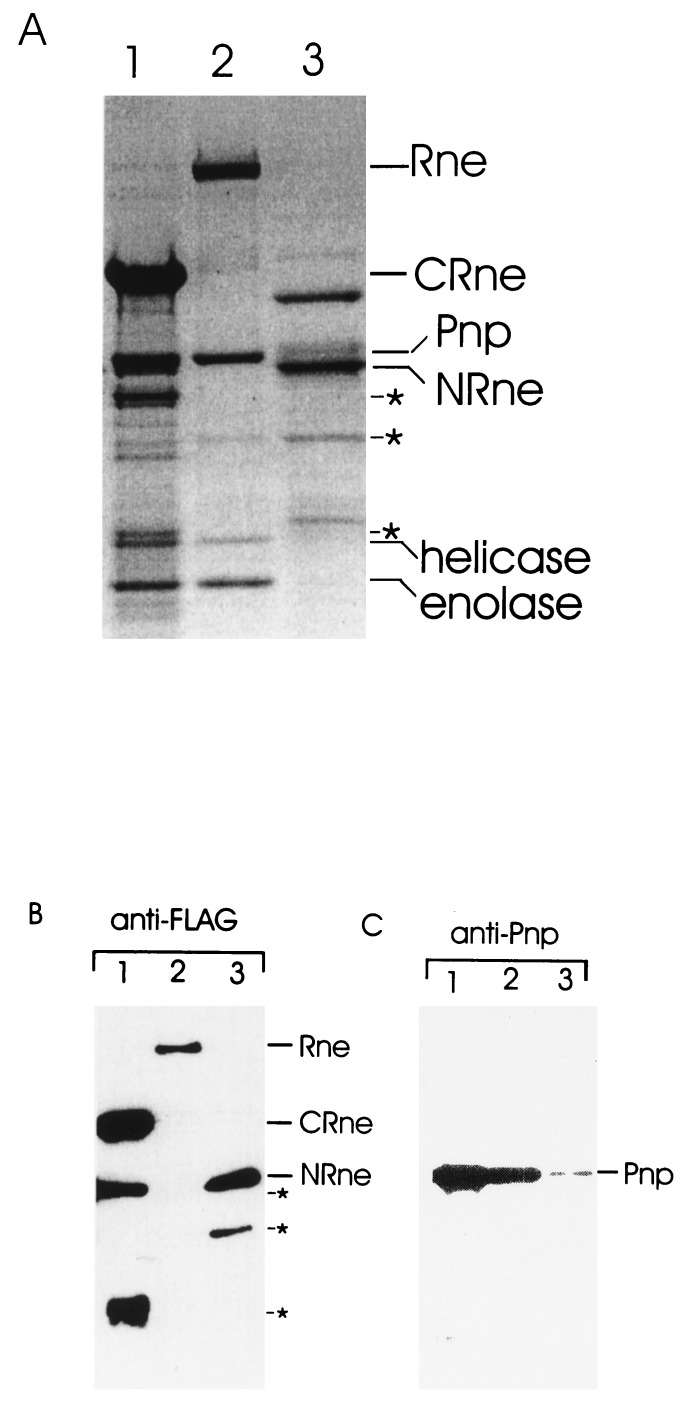
Escherichia coli RNase E, an essential single-stranded specific endoribonuclease, is required for both ribosomal RNA processing and the rapid degradation of mRNA. The availability of the complete sequences of a number of bacterial genomes prompted us to assess the evolutionarily conservation of bacterial RNase E. We show here that the sequence of the N-terminal endoribonucleolytic domain of RNase E is evolutionarily conserved in Synechocystis sp. and other bacteria. Furthermore, we demonstrate that the Synechocystis sp. homologue binds RNase E substrates and cleaves them at the same position as the E. coli enzyme. Taken together these results suggest that RNase E-mediated mechanisms of RNA decay are not confined to E. coli and its close relatives. We also show that the C-terminal half of E. coli RNase E is both sufficient and necessary for its physical interaction with the 3'-5' exoribonuclease polynucleotide phosphorylase, the RhlB helicase, and the glycolytic enzyme enolase, which are components of a "degradosome" complex. Interestingly, however, the sequence of the C-terminal half of E. coli RNase E is not highly conserved evolutionarily, suggesting diversity of RNase E interactions with other RNA decay components in different organisms. This notion is supported by our finding that the Synechocystis sp. RNase E homologue does not function as a platform for assembly of E. coli degradosome components.
Figures
References
-
- Cohen S N, McDowall K J. Mol Microbiol. 1997;23:1099–1106. - PubMed
-
- Apirion D, Lassar A B. J Biol Chem. 1978;253:1738–1742. - PubMed
-
- Ehretsmann C P, Carpousis A J, Krisch H M. FASEB J. 1992;6:3186–3192. - PubMed
-
- Melefors Ö, Lundberg U, von Gabain A. In: Control of Messenger RNA Stability. Belasco J, Brawerman G, editors. San Diego: Academic; 1993. pp. 53–70.
-
- Lin-Chao S, Cohen S N. Cell. 1991;65:1233–1242. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
