

Conformational gating of the electron transfer reaction QA-.QB --> QAQB-. in bacterial reaction centers of Rhodobacter sphaeroides determined by a driving force assay
- PMID: 9751725
- PMCID: PMC21700
- DOI: 10.1073/pnas.95.20.11679
Conformational gating of the electron transfer reaction QA-.QB --> QAQB-. in bacterial reaction centers of Rhodobacter sphaeroides determined by a driving force assay
Abstract
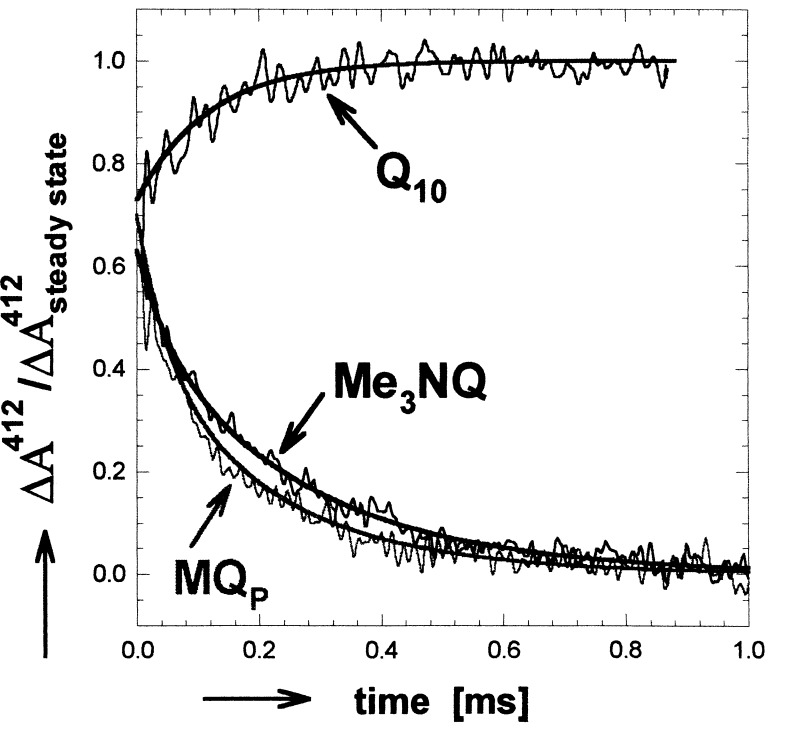
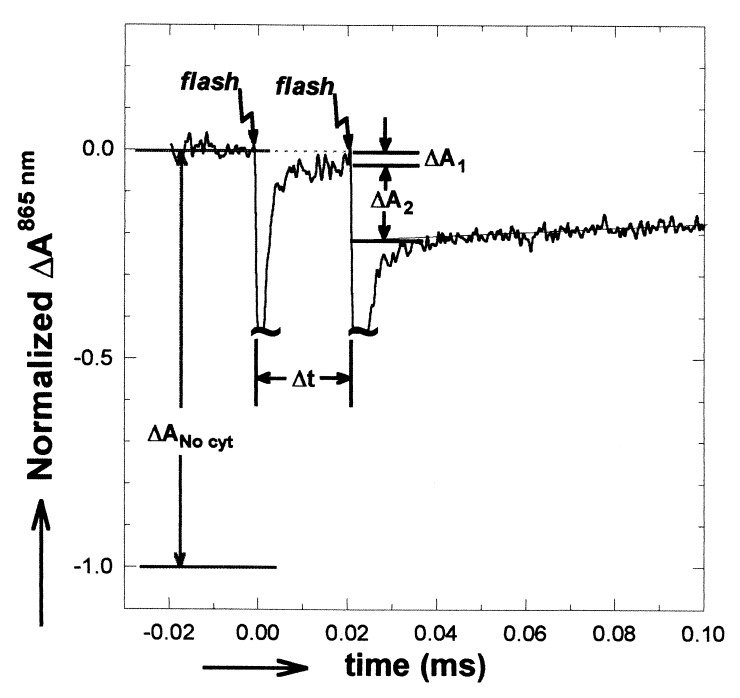
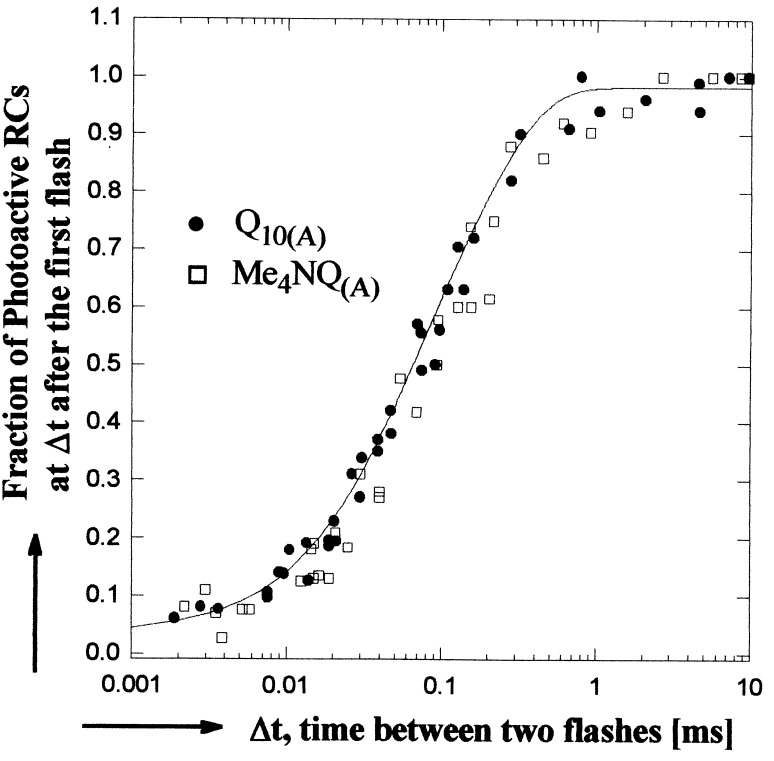
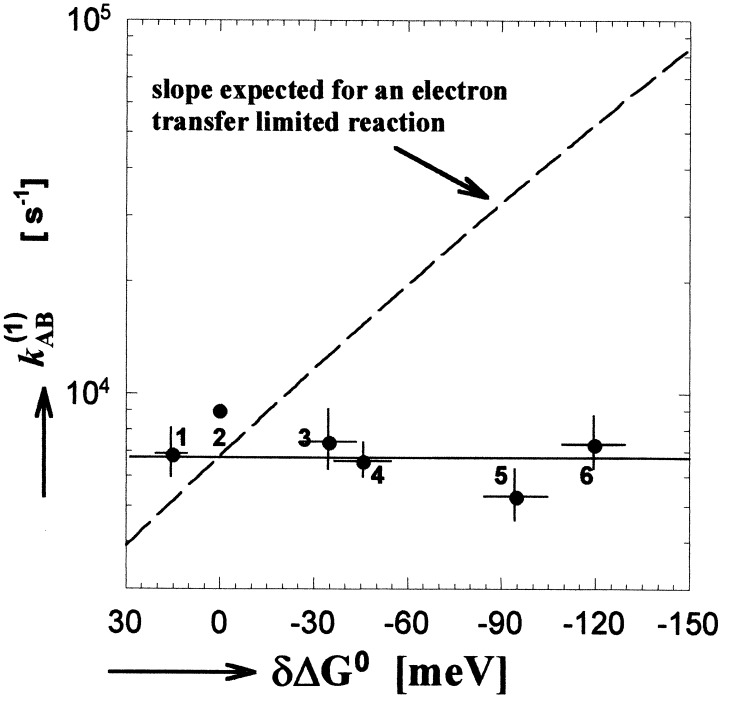
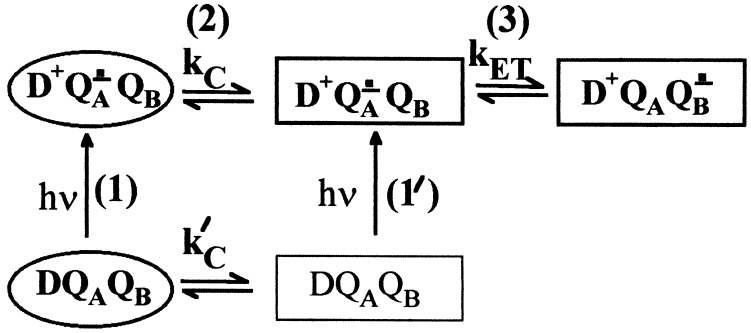
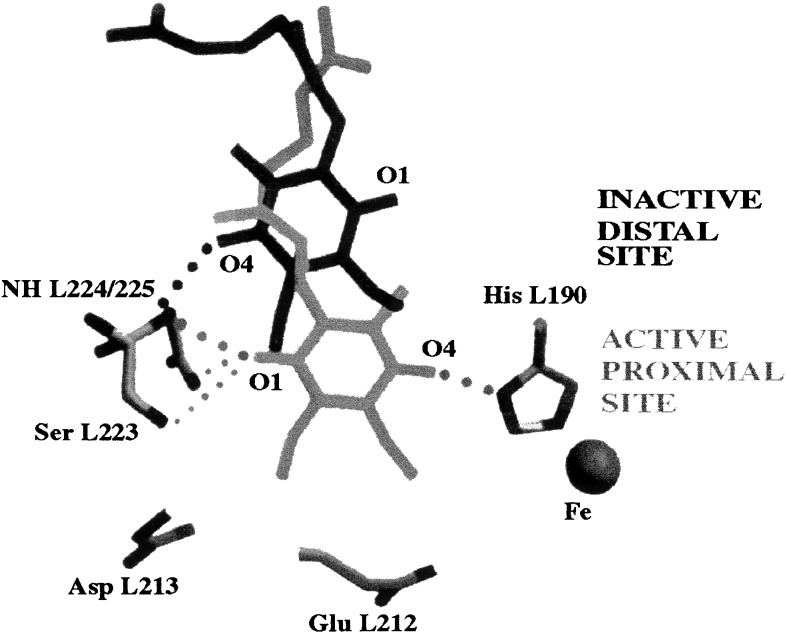
The mechanism of the electron transfer reaction, QA-.QB --> QAQB-., was studied in isolated reaction centers from the photosynthetic bacterium Rhodobacter sphaeroides by replacing the native Q10 in the QA binding site with quinones having different redox potentials. These substitutions are expected to change the intrinsic electron transfer rate by changing the redox free energy (i.e., driving force) for electron transfer without affecting other events that may be associated with the electron transfer (e.g., protein dynamics or protonation). The electron transfer from QA-. to QB was measured by three independent methods: a functional assay involving cytochrome c2 to measure the rate of QA-. oxidation, optical kinetic spectroscopy to measure changes in semiquinone absorption, and kinetic near-IR spectroscopy to measure electrochromic shifts that occur in response to electron transfer. The results show that the rate of the observed electron transfer from QA-. to QB does not change as the redox free energy for electron transfer is varied over a range of 150 meV. The strong temperature dependence of the observed rate rules out the possibility that the reaction is activationless. We conclude, therefore, that the independence of the observed rate on the driving force for electron transfer is due to conformational gating, that is, the rate limiting step is a conformational change required before electron transfer. This change is proposed to be the movement, controlled kinetically either by protein dynamics or intermolecular interactions, of QB by approximately 5 A as observed in the x-ray studies of Stowell et al. [Stowell, M. H. B., McPhillips, T. M., Rees, D. C., Soltis, S. M., Abresch, E. & Feher, G. (1997) Science 276, 812-816].
Figures
References
-
- McCammon J A, Harvey S C. Dynamics of Proteins and Nucleic Acids. New York: Cambridge Univ. Press; 1987.
-
- Hoffman B M, Ratner M A. J Am Chem Soc. 1987;109:6237–6242.
-
- Feher G, Allen J P, Okamura M Y, Rees D C. Nature (London) 1989;339:111–116.
-
- Okamura M Y, Feher G. Annu Rev Biochem. 1992;61:861–896. - PubMed
-
- Shinkarev V P, Wraight C A. In: The Photosynthetic Reaction Center. Deisenhofer J, Norris J R, editors. Vol. 1. New York: Academic; 1993. pp. 193–255.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
