

Distinct steady-state nuclear receptor coregulator complexes exist in vivo
- PMID: 9751728
- PMCID: PMC21703
- DOI: 10.1073/pnas.95.20.11697
Distinct steady-state nuclear receptor coregulator complexes exist in vivo
Abstract
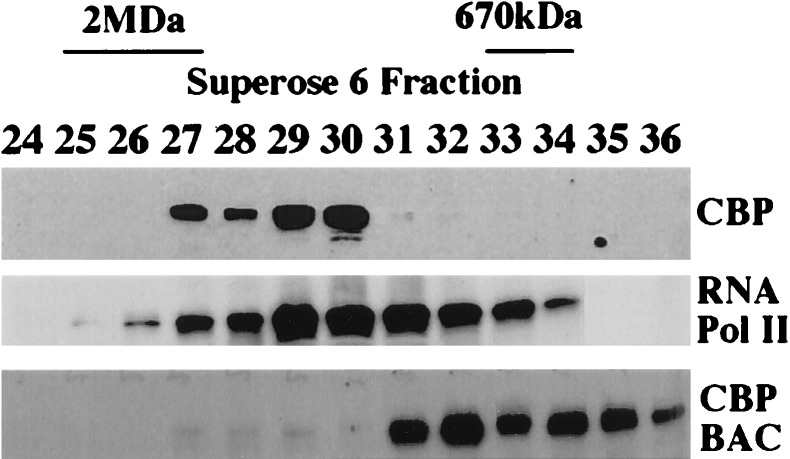
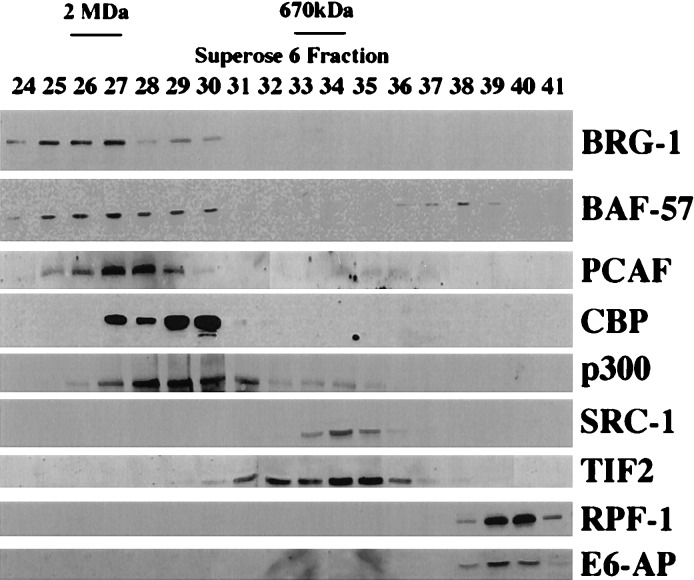
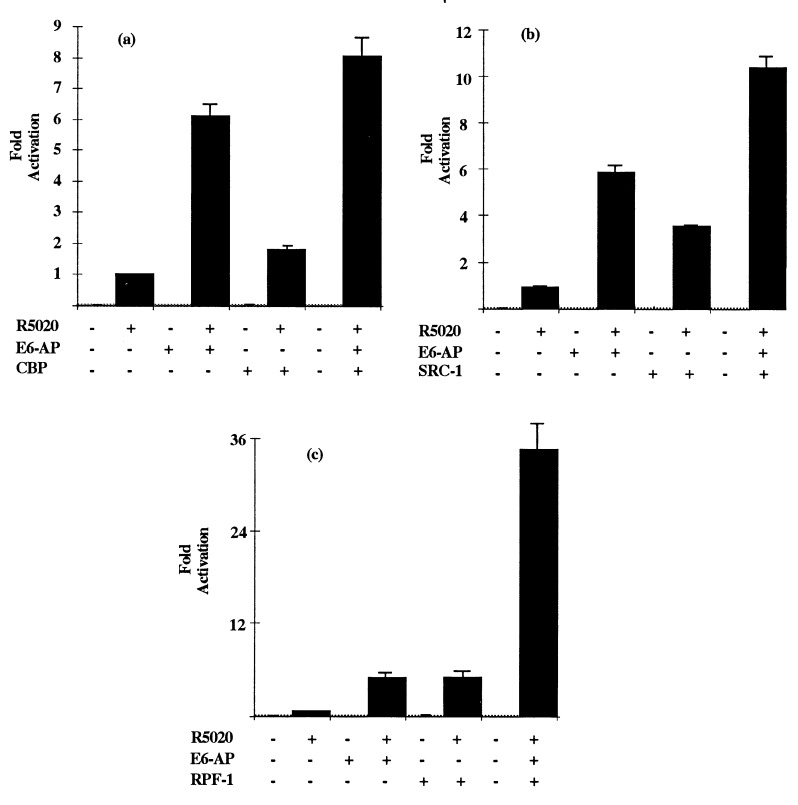
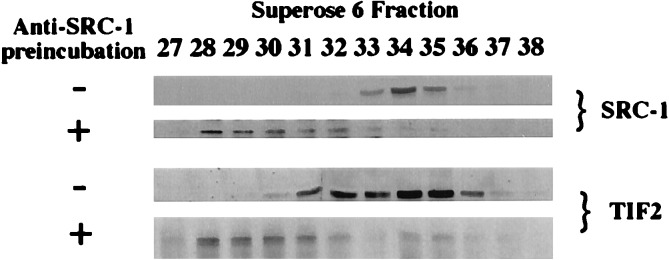
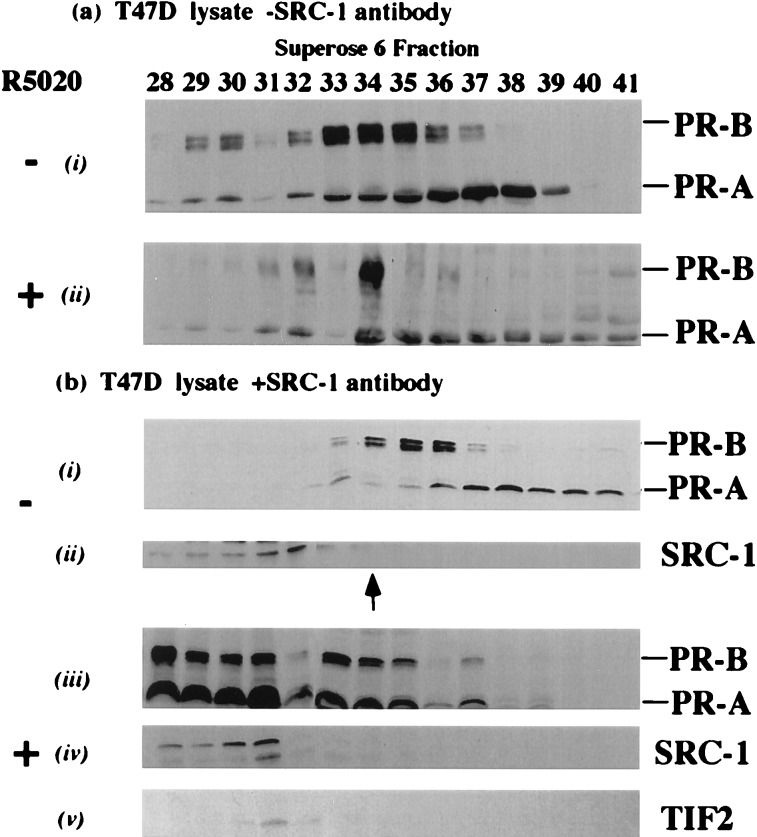
Transcriptional regulation by members of the nuclear hormone receptor superfamily is a modular process requiring the mediation of distinct subclasses of coregulators. These subclasses include members of the steroid receptor coactivator-1 (SRC-1) coactivator family, p300/CBP and their associated proteins, such as p300/CBP-associated factor, human homologs of SWI/SNF proteins such as BRG-1, and the less well-characterized E3 ubiquitin-protein ligases E6 papillomavirus protein-associated protein and receptor-potentiating factor-1. Because functional studies indicate that these coregulators may form higher order complexes, we analyzed steady-state complexes of different coregulator subclasses in vivo. T47D and HeLa cell lysates were subjected to biochemical fractionation and screened by immunoblotting using coregulator-specific antibodies. We show that different subclasses of nuclear receptor coregulators exhibit distinct fractionation profiles. Furthermore, evidence is provided that SRC-1 family members may exist in vivo in heteromultimeric forms with each other. In addition, we demonstrate that liganded PR is present in stable complexes containing SRC-1 and transcription intermediary factor 2 (TIF2) in vivo. Our results suggest that the assembly of large, modular transcriptional complexes by recruitment of distinct subclasses of preformed coregulator subcomplexes may be involved in transcriptional regulation by activated nuclear receptors.
Figures

References
-
- Tsai M-J, O’Malley B W. Annu Rev Biochem. 1994;63:451–486. - PubMed
-
- Onate S A, Tsai S Y, Tsai M-J, O’Malley B W. Science. 1995;270:1354–1357. - PubMed
-
- Kamei Y, Xu L, Heinzel T, Torchia J, Kurokawa R, Gloss B, Lin S C, Heyman R A, Rose D W, Glass C K, et al. Cell. 1996;85:403–414. - PubMed
-
- Halachmi S, Marden E, Martin G, Mackay H, Abbondanza C, Brown M. Science. 1994;264:1455–1458. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
