

Habitat structure determines competition intensity and invasion success in gecko lizards
- PMID: 9751735
- PMCID: PMC21710
- DOI: 10.1073/pnas.95.20.11739
Habitat structure determines competition intensity and invasion success in gecko lizards
Abstract
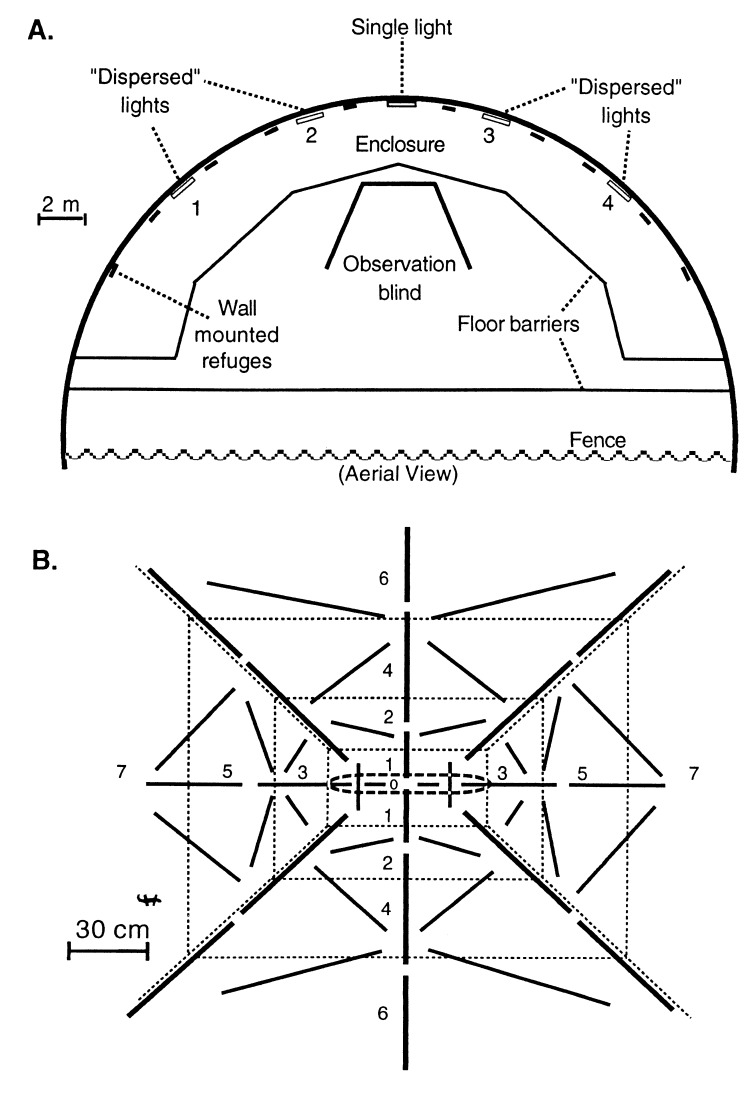
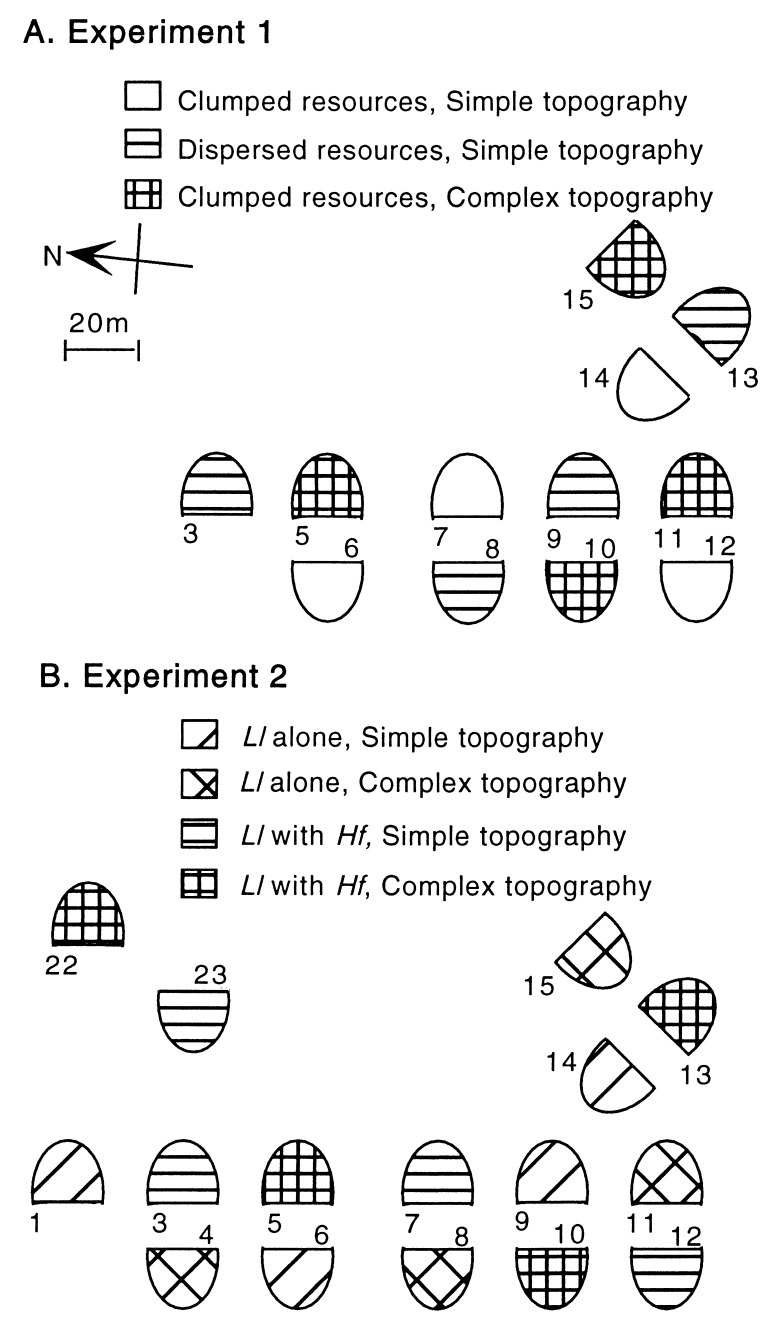
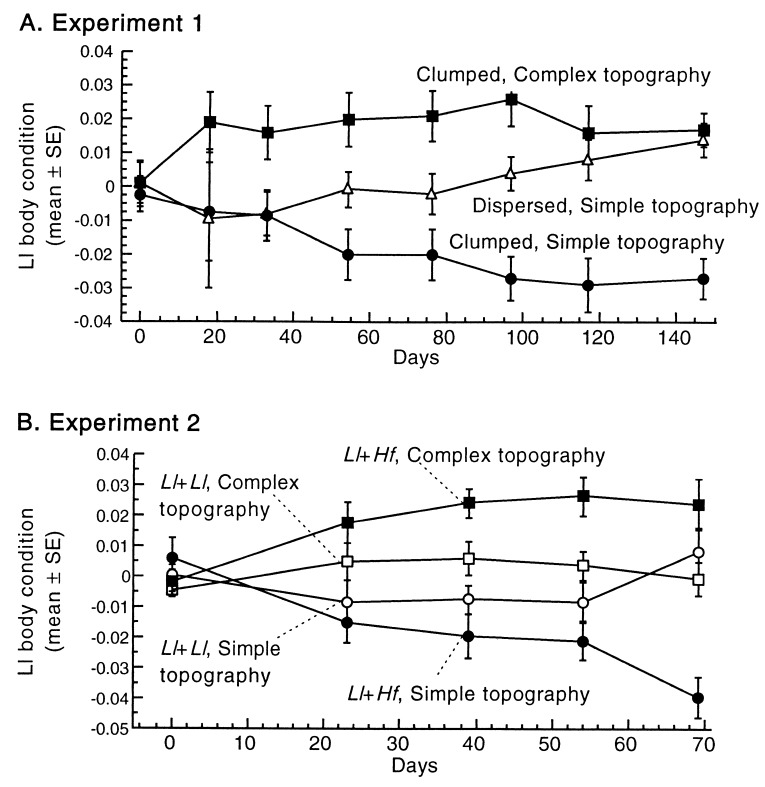
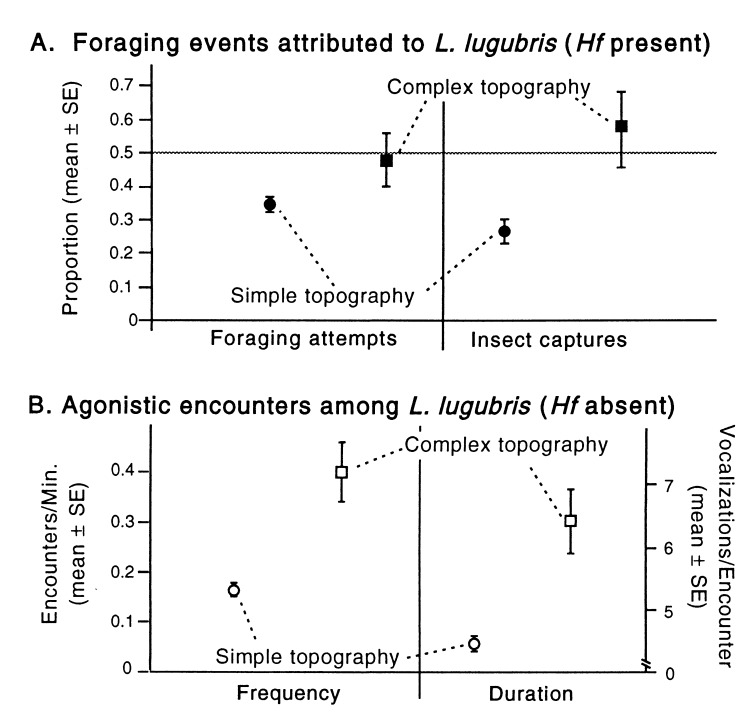
Species diversity is correlated with structural complexity in many animal communities; however, experimental tests of the mechanisms underlying this important relationship are rare, especially in terrestrial communities. We manipulated physical features of the habitat of gecko lizards and measured the effect on exploitation competition for insects. Increasing both the dispersion of food resources and microhabitat topography dramatically reduced interspecific competition. Adding topographic structure reduced the advantages of the larger, faster, invasive species. Interindividual spacing decreased, but intraspecific agonistic interference increased in the more territorial, resident species. Human structural alterations of the environment facilitate invasion and competitive displacement in this system. Physical microhabitat structure can potentially affect species interactions through a variety of complex mechanisms.
Figures

References
-
- McCoy E D, Bell S S. In: Habitat Structure: The Physical Arrangement of Objects in Space. Bell S S, McCoy E D, Mushinsky H R, editors. London: Chapman & Hall; 1991. pp. 3–27.
-
- MacArthur R H, MacArthur J W. Ecology. 1961;42:594–598.
-
- Sanders H L. Am Nat. 1968;102:243–282.
-
- Karr J R, Roth R R. Am Nat. 1971;105:423–435.
-
- Lawton J H. Annu Rev Entomol. 1983;28:23–39.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
