

Protein rearrangements underlying slow inactivation of the Shaker K+ channel
- PMID: 9758858
- PMCID: PMC2229423
- DOI: 10.1085/jgp.112.4.377
Protein rearrangements underlying slow inactivation of the Shaker K+ channel
Abstract
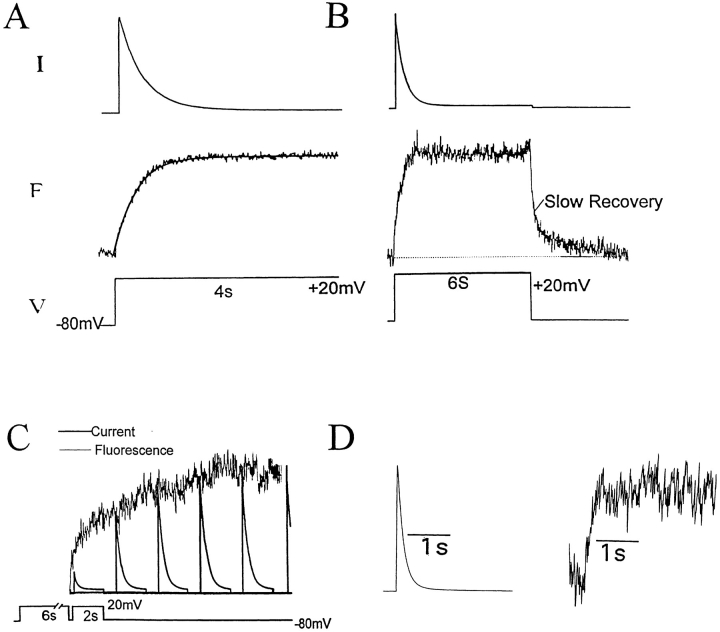
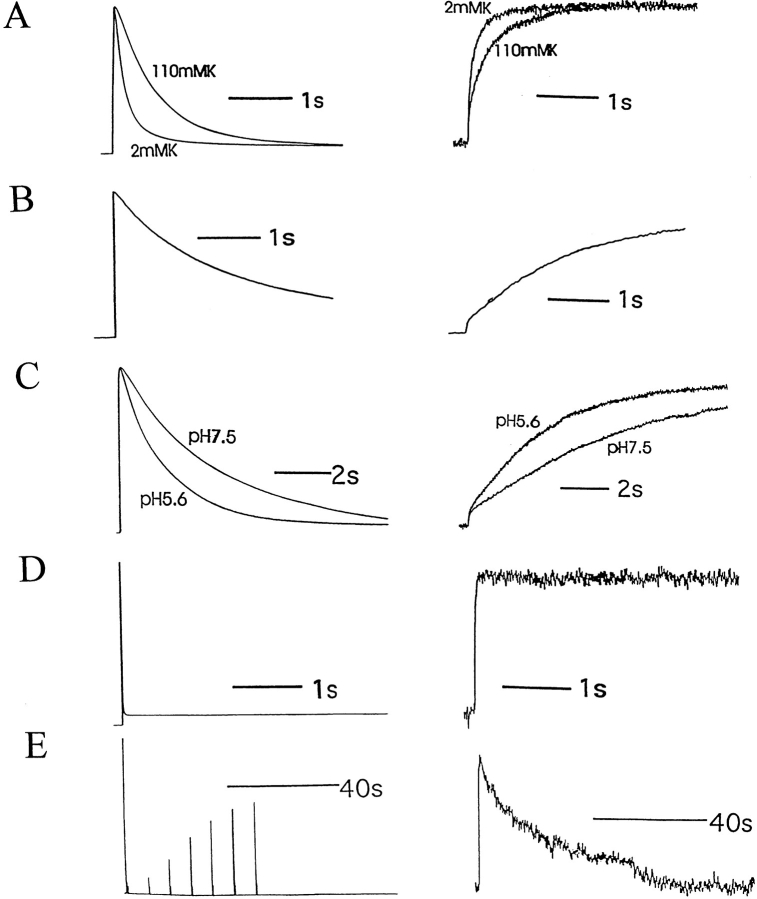
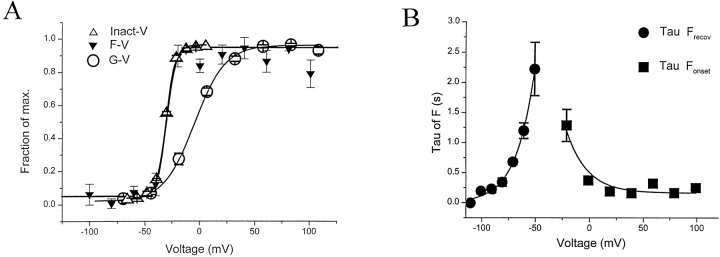
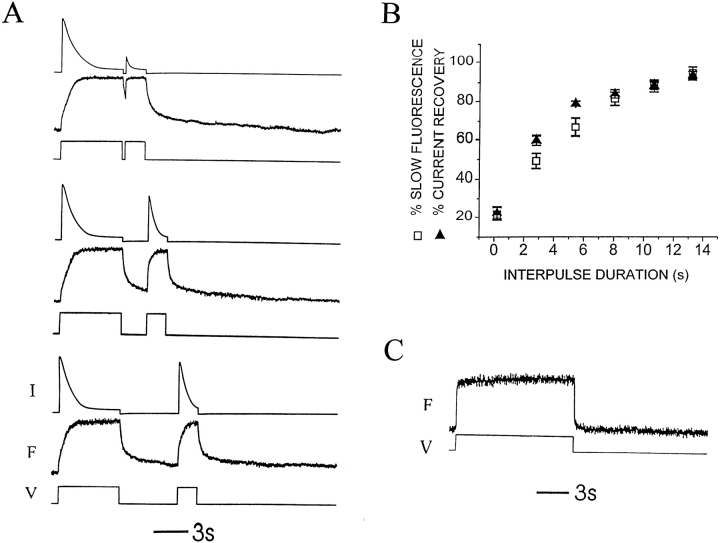
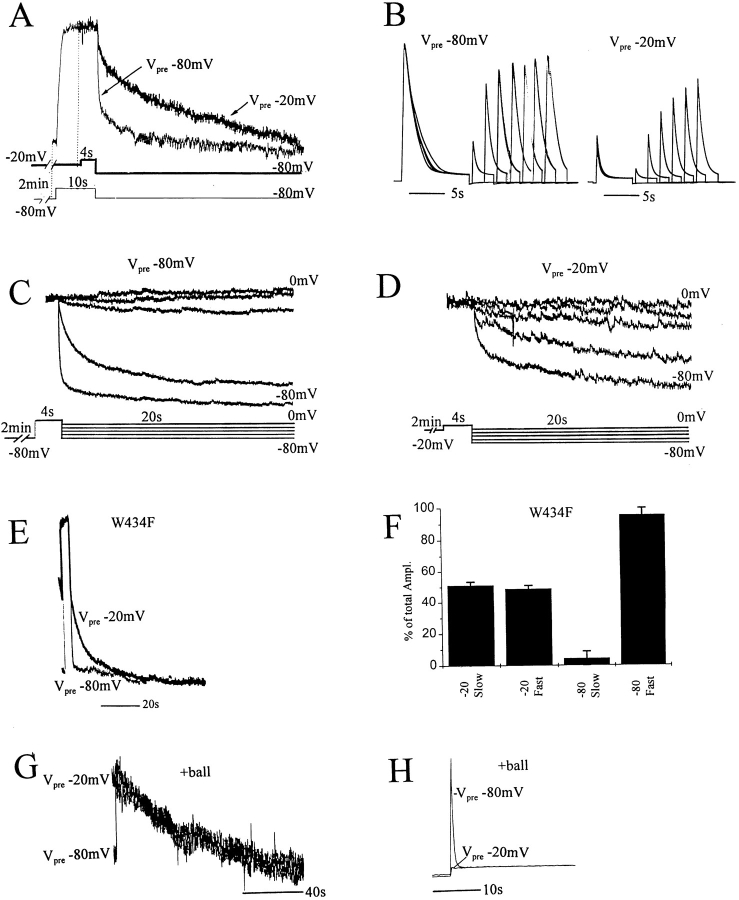
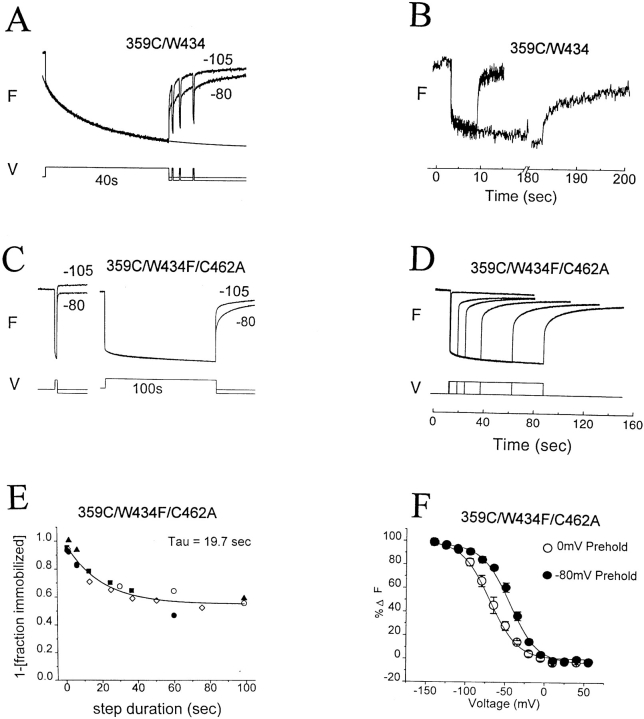
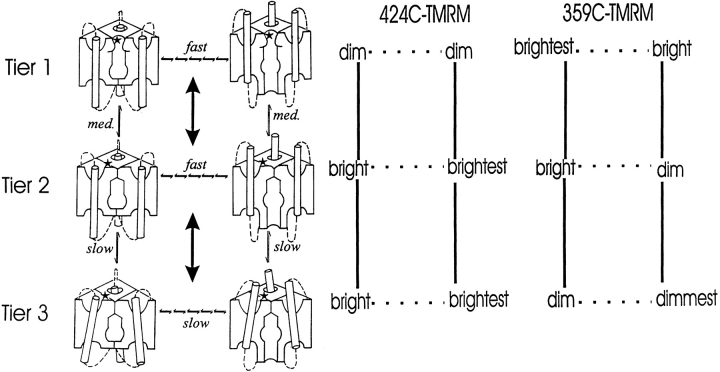
Voltage-dependent ion channels transduce changes in the membrane electric field into protein rearrangements that gate their transmembrane ion permeation pathways. While certain molecular elements of the voltage sensor and gates have been identified, little is known about either the nature of their conformational rearrangements or about how the voltage sensor is coupled to the gates. We used voltage clamp fluorometry to examine the voltage sensor (S4) and pore region (P-region) protein motions that underlie the slow inactivation of the Shaker K+ channel. Fluorescent probes in both the P-region and S4 changed emission intensity in parallel with the onset and recovery of slow inactivation, indicative of local protein rearrangements in this gating process. Two sequential rearrangements were observed, with channels first entering the P-type, and then the C-type inactivated state. These forms of inactivation appear to be mediated by a single gate, with P-type inactivation closing the gate and C-type inactivation stabilizing the gate's closed conformation. Such a stabilization was due, at least in part, to a slow rearrangement around S4 that stabilizes S4 in its activated transmembrane position. The fluorescence reports of S4 and P-region fluorophore are consistent with an increased interaction of the voltage sensor and inactivation gate upon gate closure, offering insight into how the voltage-sensing apparatus is coupled to a channel gate.
Figures
References
-
- Baker OS, Larsson HP, Mannuzzu LM, Isacoff EY. Three transmembrane conformations and sequence-dependent displacement of the S4 domain in Shaker K+channel gating. Neuron. 1998;20:1283–1294. - PubMed
-
- Baukrowitz T, Yellen G. Modulation of K+ current by frequency and external [K+]: a tale of two inactivation mechanisms. Neuron. 1995;15:951–960. - PubMed
-
- Baukrowitz T, Yellen G. Use-dependent blockers and exit rate of the last ion from the multi-ion pore of a K+channel. Science. 1996;271:653–656. - PubMed
-
- Bezanilla F, Perozo E, Papazian D, Stefani E. Molecular basis of gating charge immobilization in Shakerpotassium channels. Science. 1991;254:679–683. - PubMed
