

Fab1p is essential for PtdIns(3)P 5-kinase activity and the maintenance of vacuolar size and membrane homeostasis
- PMID: 9763421
- PMCID: PMC2132800
- DOI: 10.1083/jcb.143.1.65
Fab1p is essential for PtdIns(3)P 5-kinase activity and the maintenance of vacuolar size and membrane homeostasis
Abstract
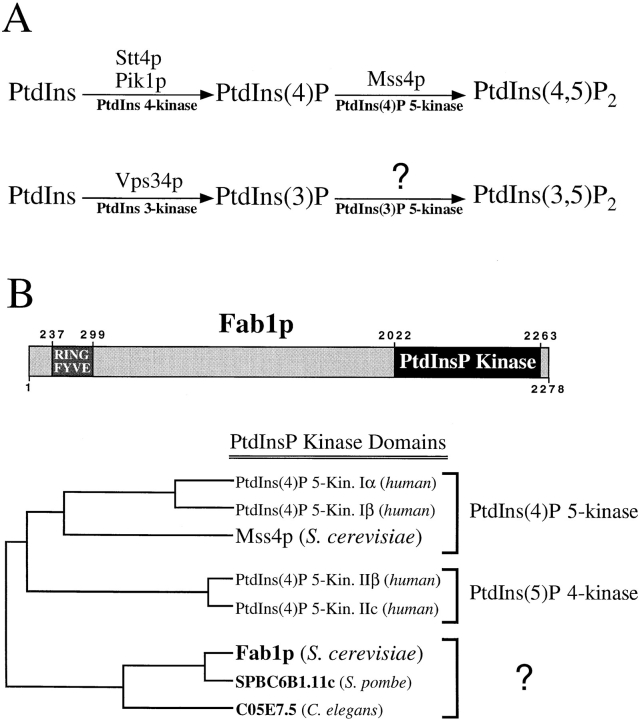
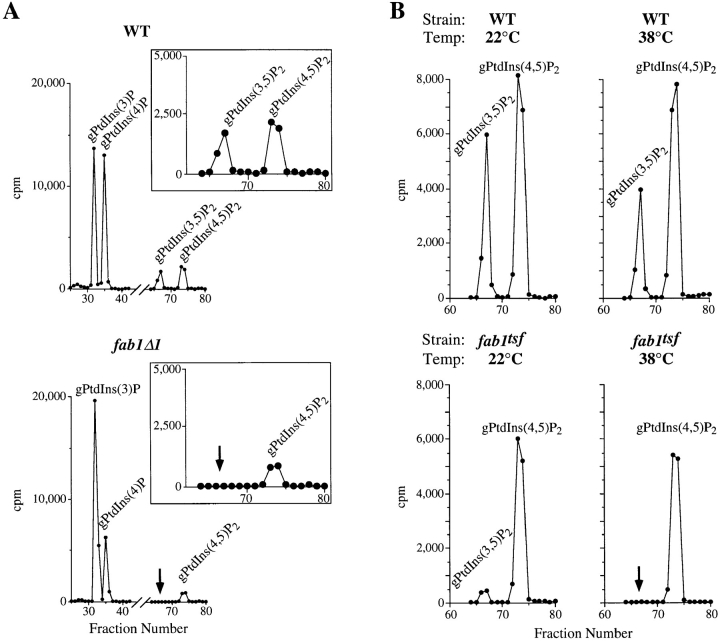
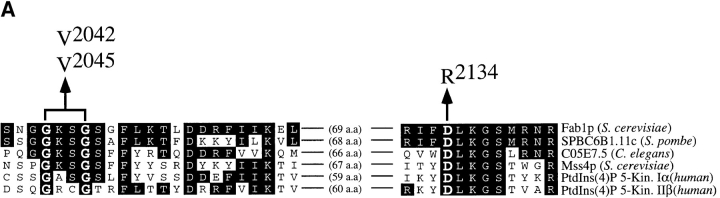
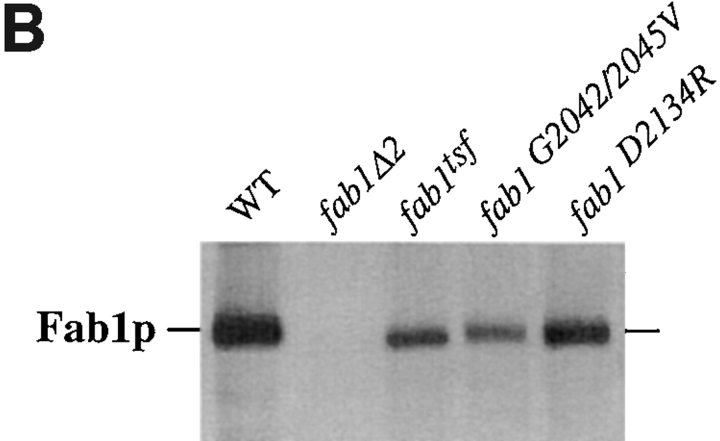
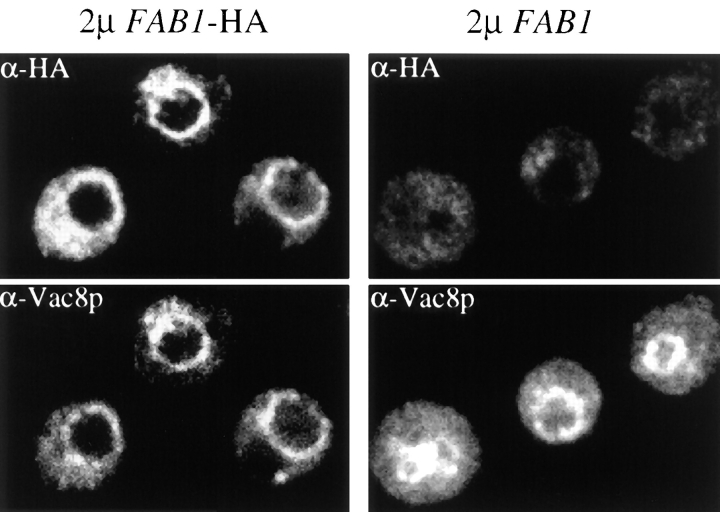
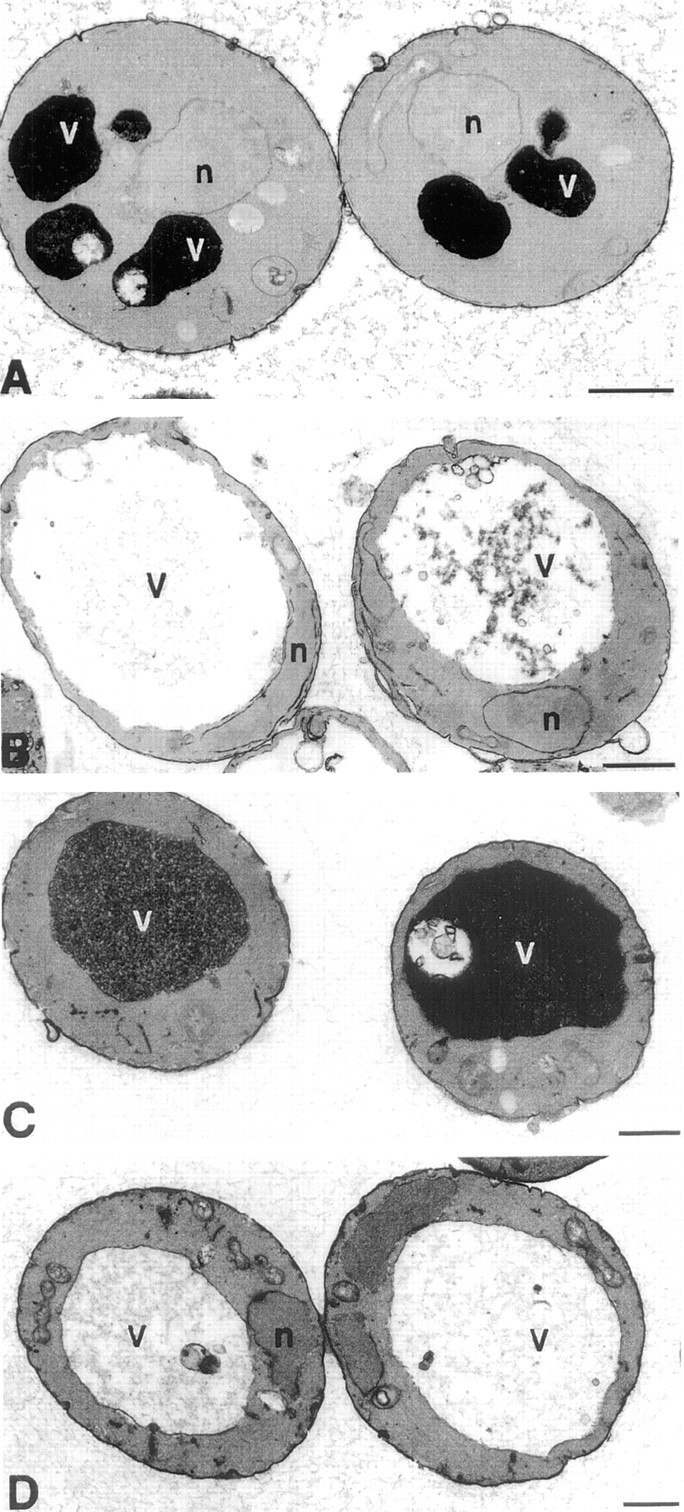
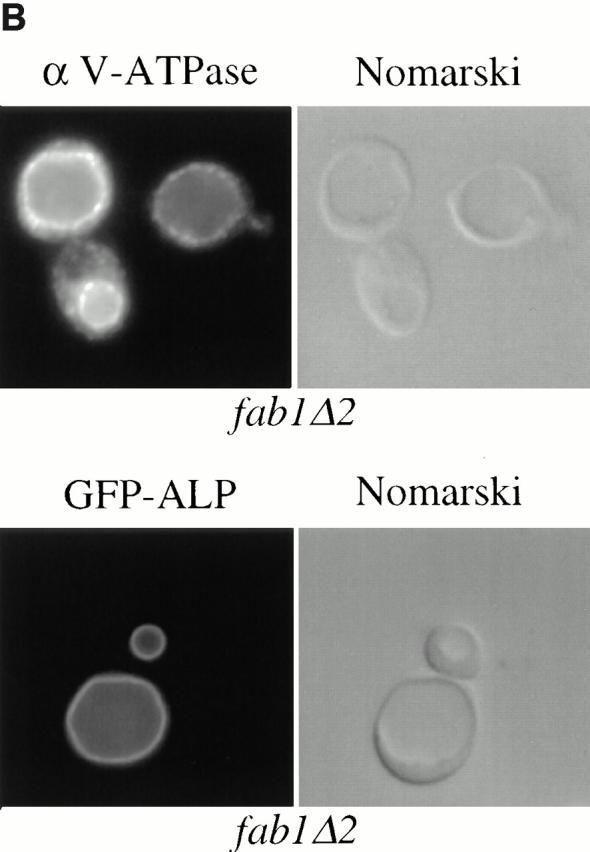
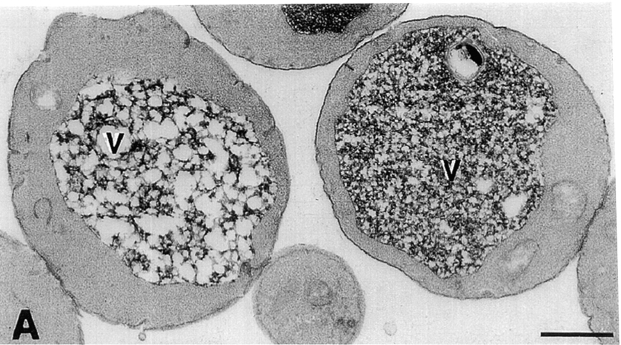
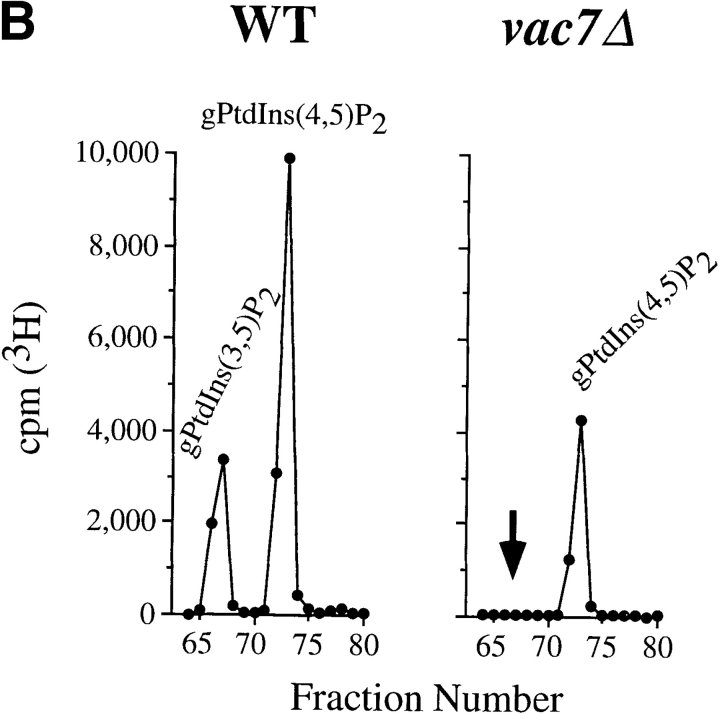
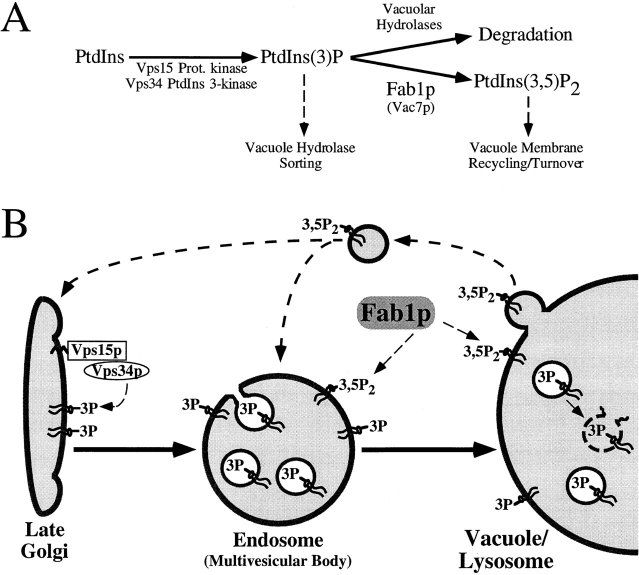
The Saccharomyces cerevisiae FAB1 gene encodes a 257-kD protein that contains a cysteine-rich RING-FYVE domain at its NH2-terminus and a kinase domain at its COOH terminus. Based on its sequence, Fab1p was initially proposed to function as a phosphatidylinositol 4-phosphate (PtdIns(4)P) 5-kinase (). Additional sequence analysis of the Fab1p kinase domain, reveals that Fab1p defines a subfamily of putative PtdInsP kinases that is distinct from the kinases that synthesize PtdIns(4,5)P2. Consistent with this, we find that unlike wild-type cells, fab1Delta, fab1(tsf), and fab1 kinase domain point mutants lack detectable levels of PtdIns(3,5)P2, a phosphoinositide recently identified both in yeast and mammalian cells. PtdIns(4,5)P2 synthesis, on the other hand, is only moderately affected even in fab1Delta mutants. The presence of PtdIns(3)P in fab1 mutants, combined with previous data, indicate that PtdIns(3,5)P2 synthesis is a two step process, requiring the production of PtdIns(3)P by the Vps34p PtdIns 3-kinase and the subsequent Fab1p- dependent phosphorylation of PtdIns(3)P yielding PtdIns(3,5)P2. Although Vps34p-mediated synthesis of PtdIns(3)P is required for the proper sorting of hydrolases from the Golgi to the vacuole, the production of PtdIns(3,5)P2 by Fab1p does not directly affect Golgi to vacuole trafficking, suggesting that PtdIns(3,5)P2 has a distinct function. The major phenotypes resulting from Fab1p kinase inactivation include temperature-sensitive growth, vacuolar acidification defects, and dramatic increases in vacuolar size. Based on our studies, we hypothesize that whereas Vps34p is essential for anterograde trafficking of membrane and protein cargoes to the vacuole, Fab1p may play an important compensatory role in the recycling/turnover of membranes deposited at the vacuole. Interestingly, deletion of VAC7 also results in an enlarged vacuole morphology and has no detectable PtdIns(3,5)P2, suggesting that Vac7p functions as an upstream regulator, perhaps in a complex with Fab1p. We propose that Fab1p and Vac7p are components of a signal transduction pathway which functions to regulate the efflux or turnover of vacuolar membranes through the regulated production of PtdIns(3,5)P2.
Figures



References
-
- Auger KR, Carpenter CL, Cantley LC, Varticovski L. Phosphatidylinositol 3-kinase and its novel product, phosphatidylinositol 3-phosphate, are present in Saccharomyces cerevisiae. . J Biol Chem. 1989;264:20181–20184. - PubMed
-
- Bankaitis VA, Aitken JR, Cleves AE, Dowhan W. An essential role for a phospholipid transfer protein in yeast Golgi function. Nature. 1990;347:561–562. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
