

Neurofilament-dependent radial growth of motor axons and axonal organization of neurofilaments does not require the neurofilament heavy subunit (NF-H) or its phosphorylation
- PMID: 9763429
- PMCID: PMC2132801
- DOI: 10.1083/jcb.143.1.171
Neurofilament-dependent radial growth of motor axons and axonal organization of neurofilaments does not require the neurofilament heavy subunit (NF-H) or its phosphorylation
Abstract
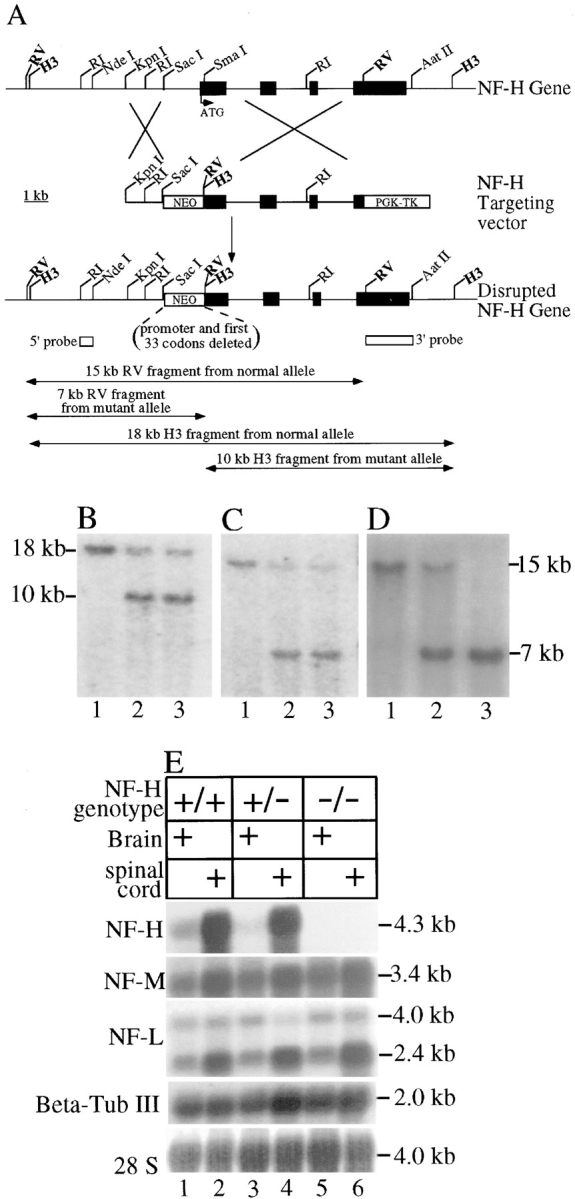
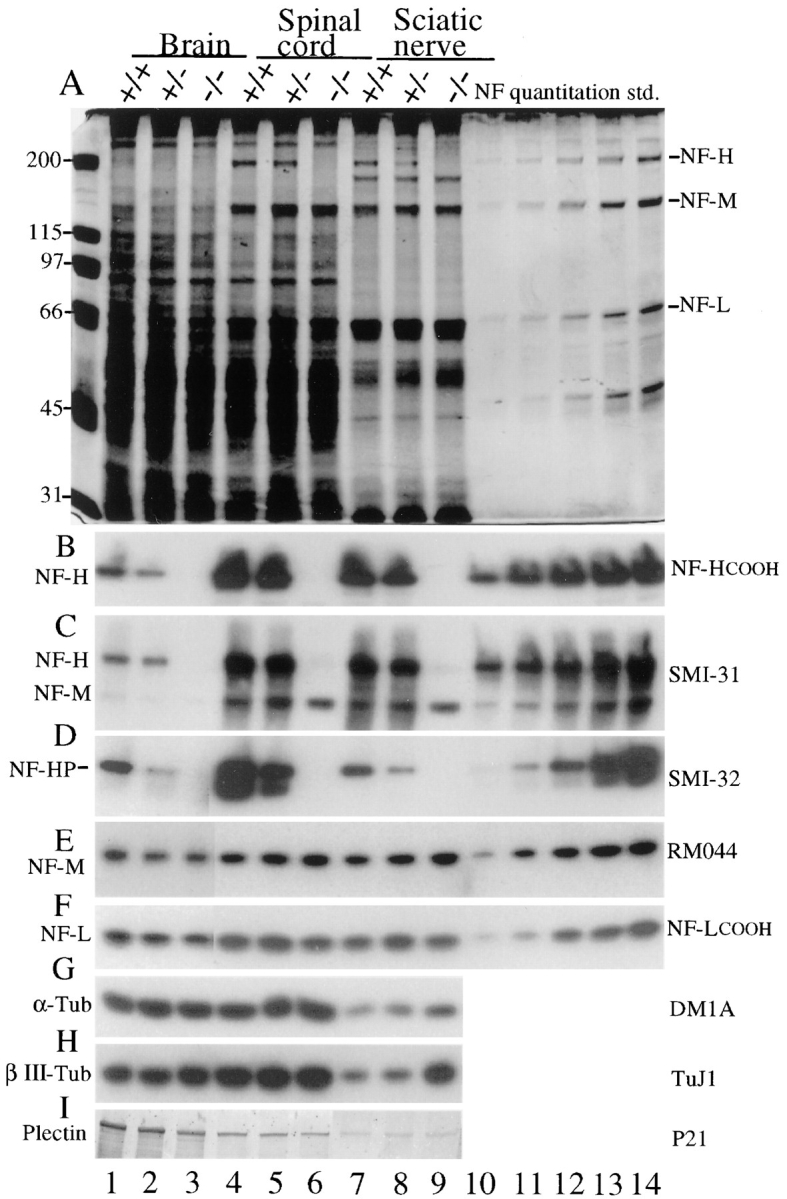
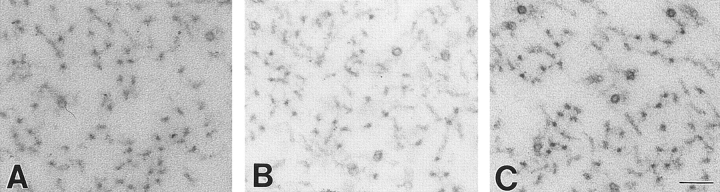
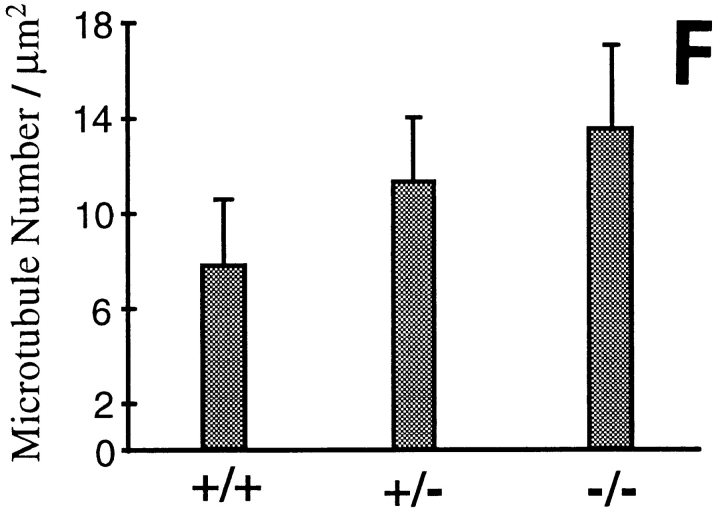
Neurofilaments are essential for establishment and maintenance of axonal diameter of large myelinated axons, a property that determines the velocity of electrical signal conduction. One prominent model for how neurofilaments specify axonal growth is that the 660-amino acid, heavily phosphorylated tail domain of neurofilament heavy subunit (NF-H) is responsible for neurofilament-dependent structuring of axoplasm through intra-axonal crossbridging between adjacent neurofilaments or to other axonal structures. To test such a role, homologous recombination was used to generate NF-H-null mice. In peripheral motor and sensory axons, absence of NF-H does not significantly affect the number of neurofilaments or axonal elongation or targeting, but it does affect the efficiency of survival of motor and sensory axons. Loss of NF-H caused only a slight reduction in nearest neighbor spacing of neurofilaments and did not affect neurofilament distribution in either large- or small-diameter motor axons. Since postnatal growth of motor axon caliber continues largely unabated in the absence of NF-H, neither interactions mediated by NF-H nor the extensive phosphorylation of it within myelinated axonal segments are essential features of this growth.
Figures





Comment in
-
Gene targeting studies begin to reveal the function of neurofilament proteins.J Cell Biol. 1998 Oct 5;143(1):1-4. doi: 10.1083/jcb.143.1.1. J Cell Biol. 1998. PMID: 9763415 Free PMC article. Review. No abstract available.
References
-
- Brown A, Bernier G, Mathieu M, Rossant J, Kothary R. The mouse dystonia musculorum gene is a neural isoform of bullous pemphigoid antigen 1. Nat Genet. 1995;10:301–306. - PubMed
-
- Carden MJ, Schlaepfer WW, Lee VM. The structure, biochemical properties, and immunogenicity of neurofilament peripheral regions are determined by phosphorylation state. J Biol Chem. 1985;260:9805–9817. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
