

Synaptic regulation of action potential timing in neostriatal cholinergic interneurons
- PMID: 9763496
- PMCID: PMC6792851
- DOI: 10.1523/JNEUROSCI.18-20-08539.1998
Synaptic regulation of action potential timing in neostriatal cholinergic interneurons
Abstract
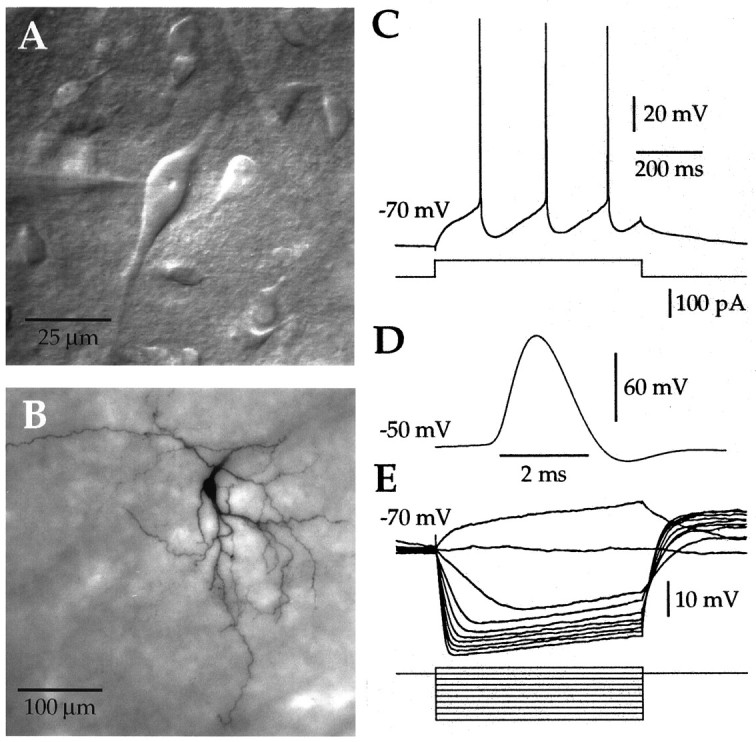
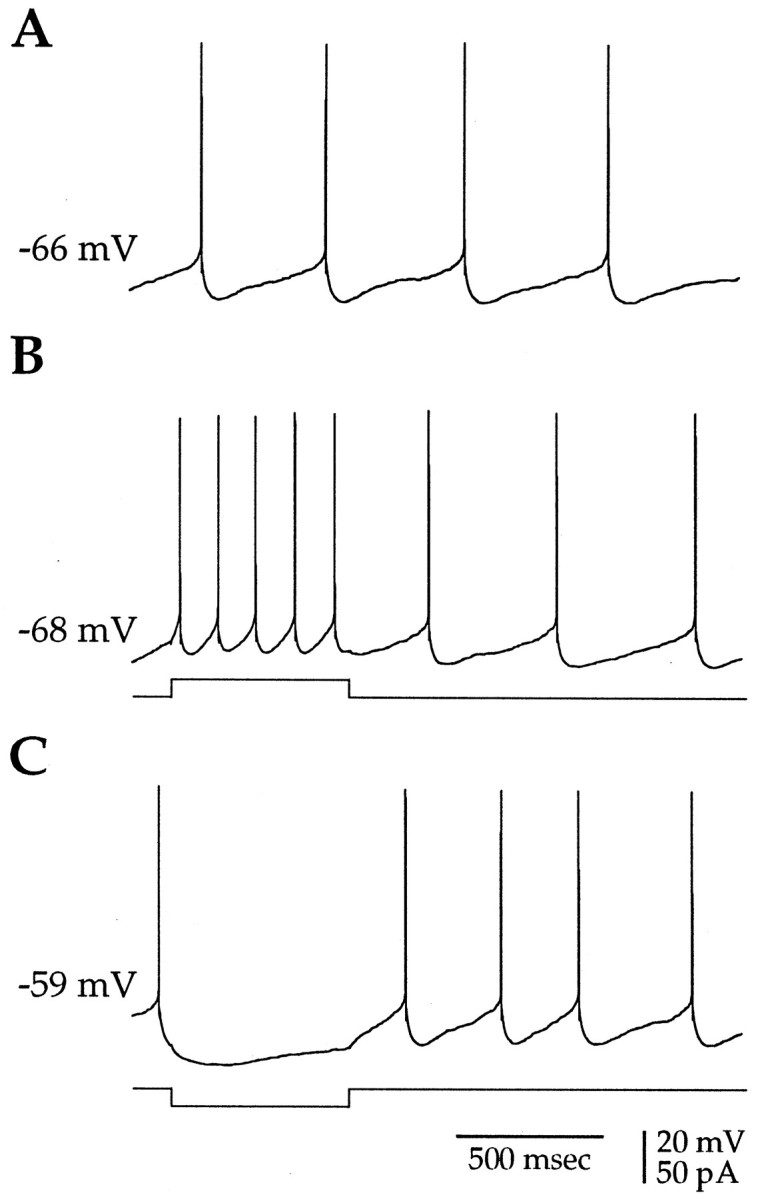
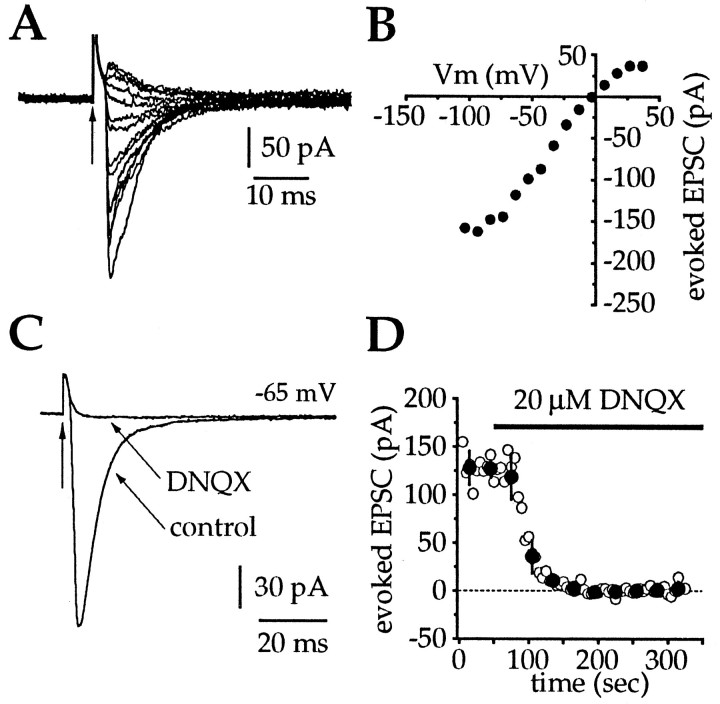
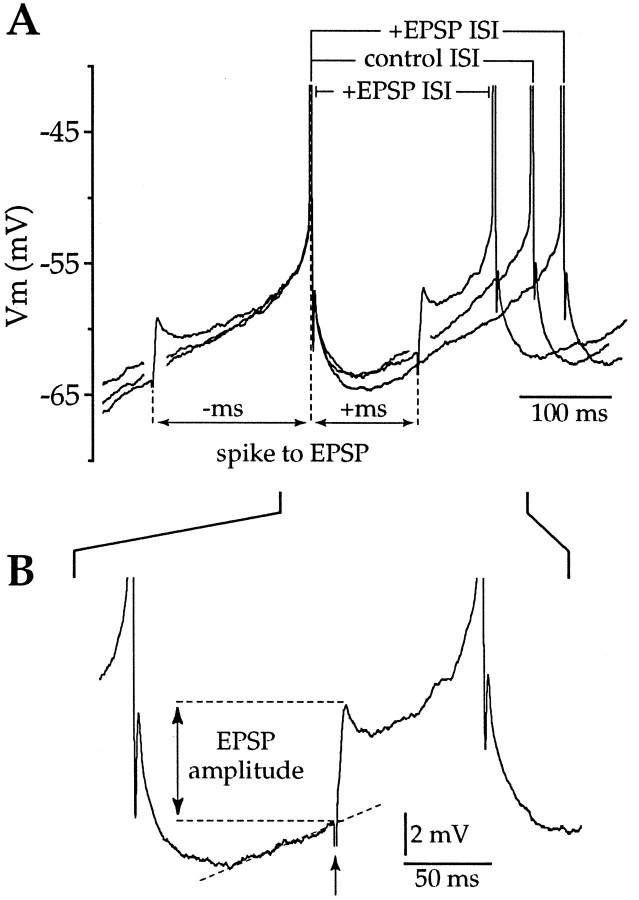
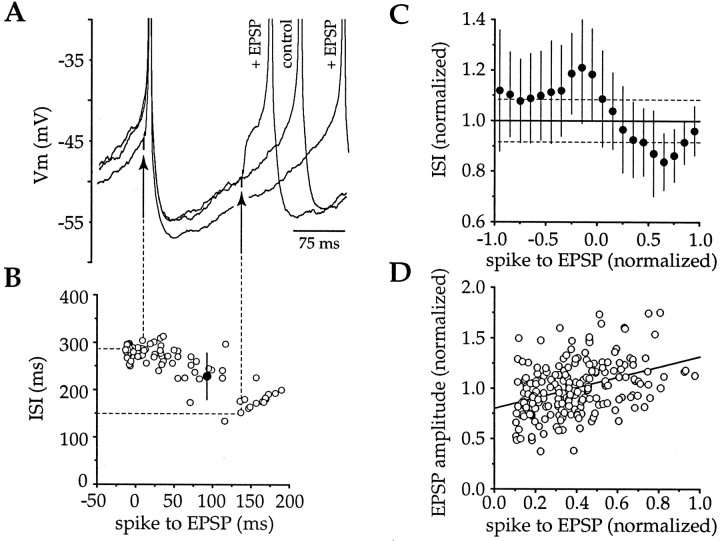
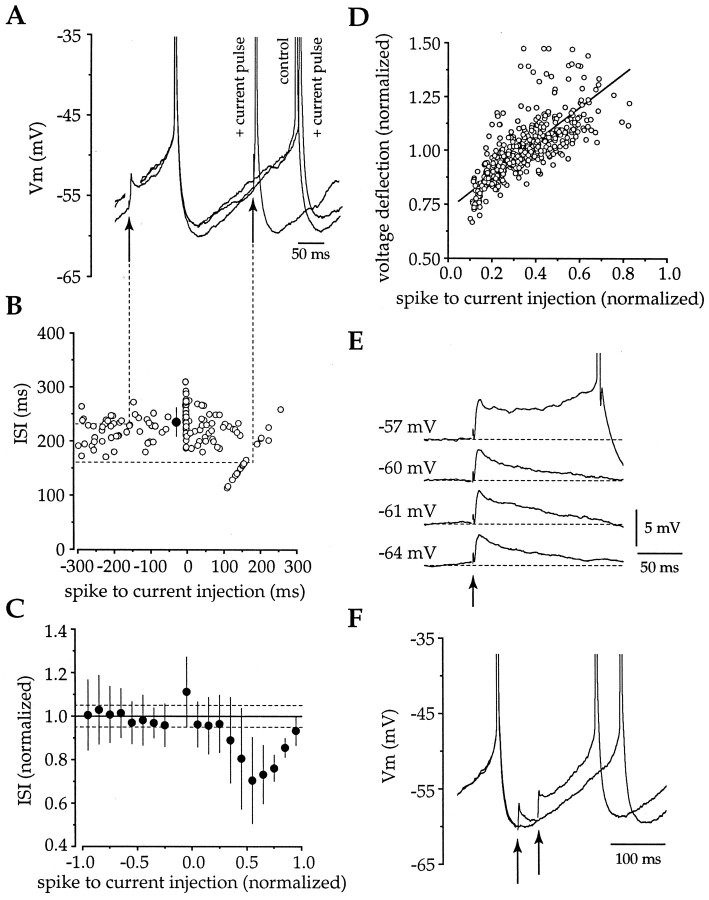
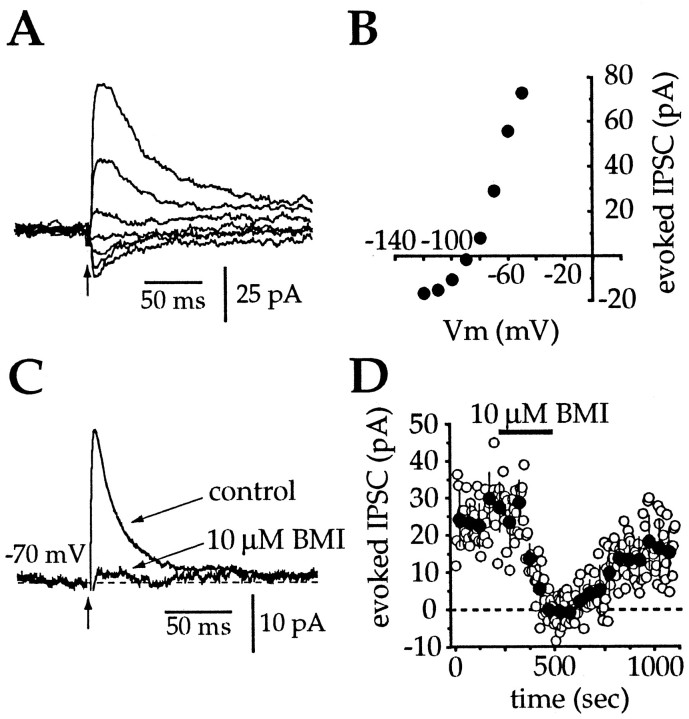
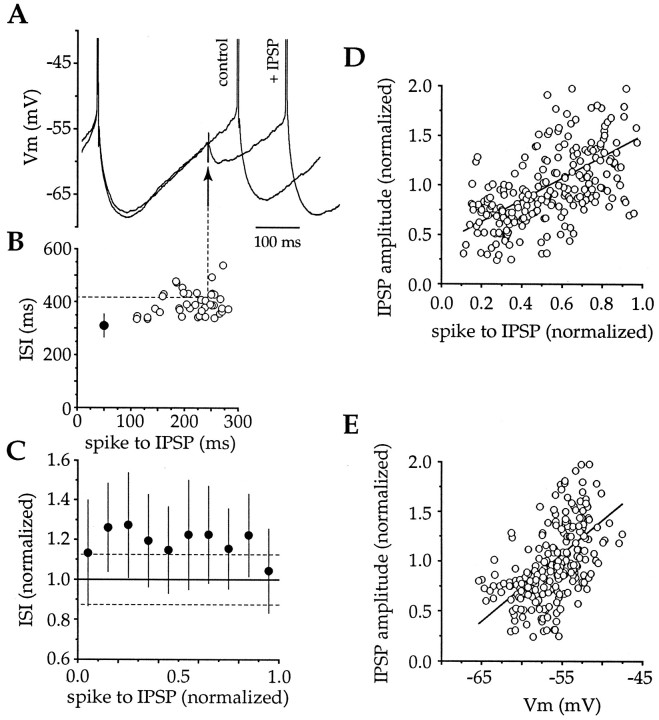
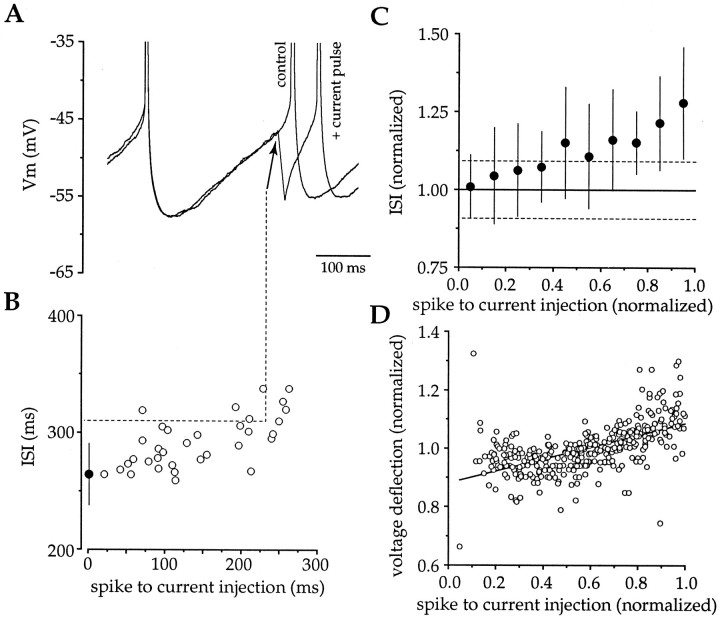
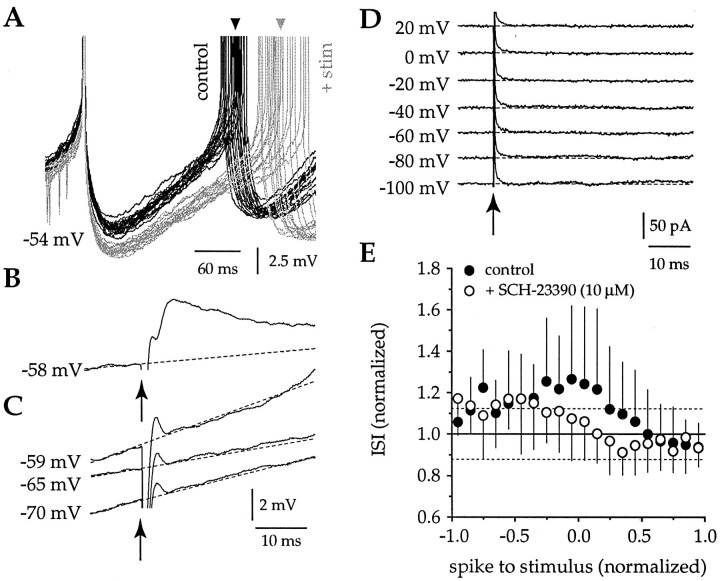
Action potentials in neostriatal cholinergic interneurons recorded in vivo are triggered by summation of two or three discrete synaptic depolarizations (Wilson et al., 1990). The ability and precision with which EPSPs and IPSPs regulate action potential timing was therefore investigated in vitro. Cholinergic interneurons were identified on the basis of morphological and electrophysiological characteristics in neostriatal slices taken from 2- to 3-week-old postnatal rats recorded at 24-26 degreesC. During periods of induced regular firing, intrastriatal stimuli were used to evoke pharmacologically isolated monosynaptic AMPA receptor-mediated EPSPs or GABAA receptor-mediated IPSPs. EPSPs evoked during the interspike interval (ISI) produced a phase-dependent decrease in the ISI, whereas IPSPs produced a phase-independent prolongation of the ISI. Injection of brief depolarizing currents mimicked the action of EPSPs and revealed an alteration in the input resistance during the ISI. In contrast to IPSPs, the ability of brief hyperpolarizing current injections to delay spike generation was phase-dependent. After blockade of GABAergic and glutamatergic synaptic transmission, stimuli failed to produce a detectable conductance change but could still prolong the subsequent ISI primarily through a D1 dopamine receptor-mediated enhancement of the afterhyperpolarization (AHP). Hence, EPSPs are ideally suited to provide a precise regulation of spike timing in cholinergic cells, whereas IPSPs are more likely to influence the overall level of excitability. The D1-mediated modulation of the AHP may contribute to the prolonged ISI seen in tonically active neurons in vivo in monkeys trained to respond to a sensory cue.
Figures
References
-
- Agard DA, Hiraoka Y, Shaw P, Sedat JW. Fluorescence microscopy in three dimensions. Methods Cell Biol. 1989;30:353–377. - PubMed
-
- Aosaki T, Graybiel AM, Kimura M. Effect of the nigrostriatal dopamine system on acquired neural responses in the striatum of behaving monkeys. Science. 1994a;265:412–415. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources