

Regulation of anti-DNA B cells in recombination-activating gene-deficient mice
- PMID: 9763604
- PMCID: PMC2212494
- DOI: 10.1084/jem.188.7.1247
Regulation of anti-DNA B cells in recombination-activating gene-deficient mice
Abstract
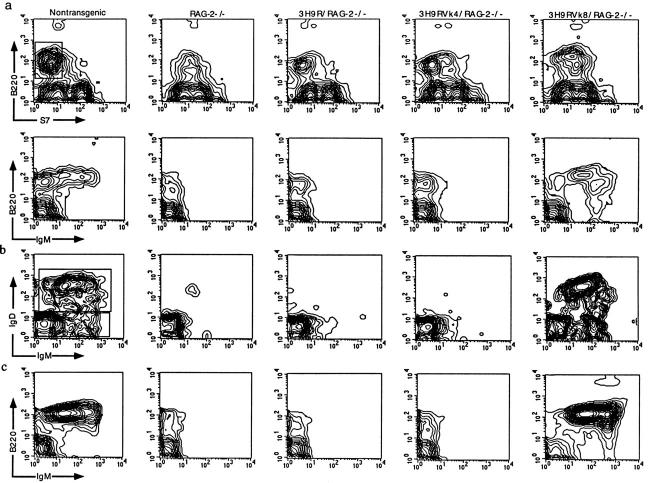
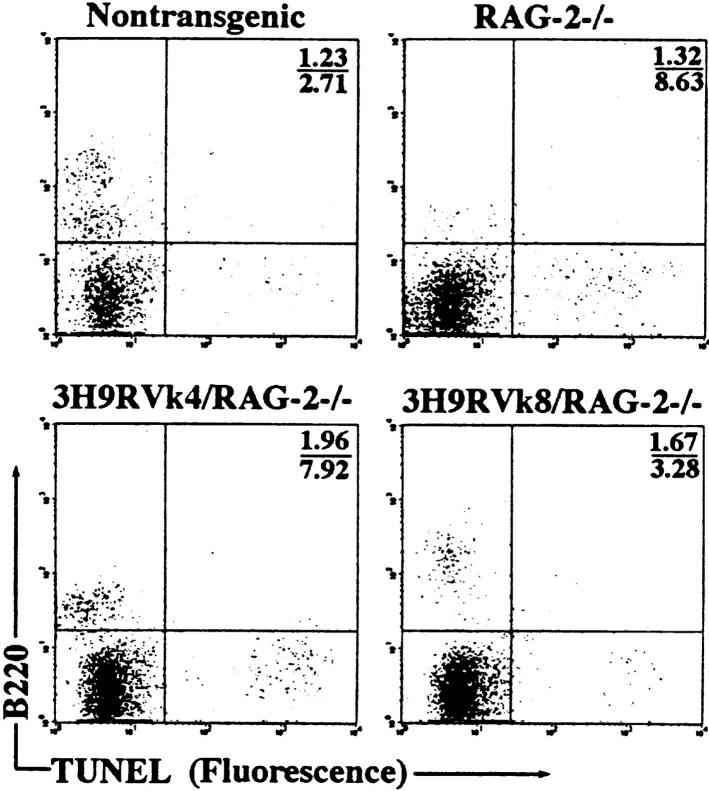
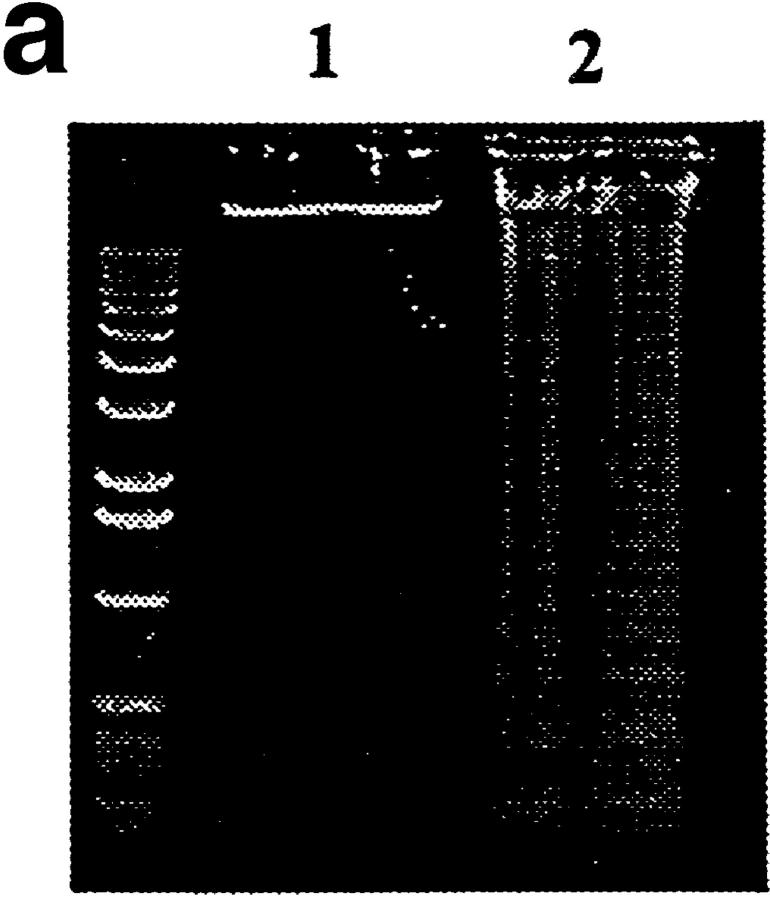
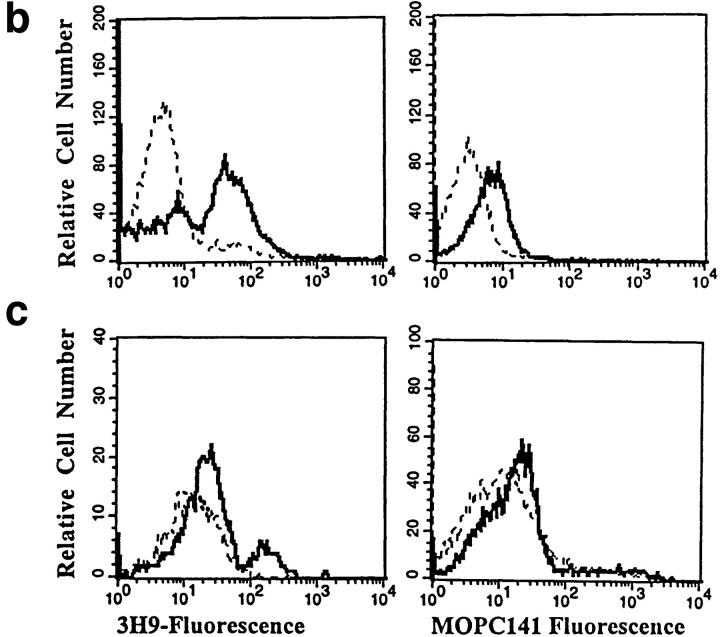
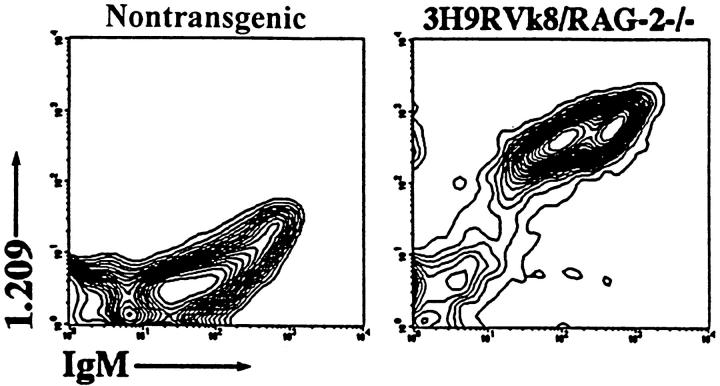
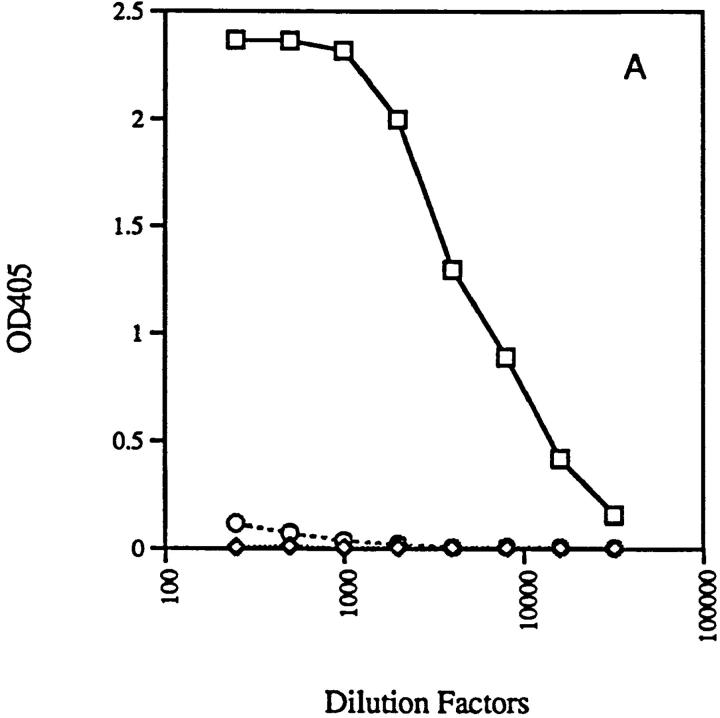
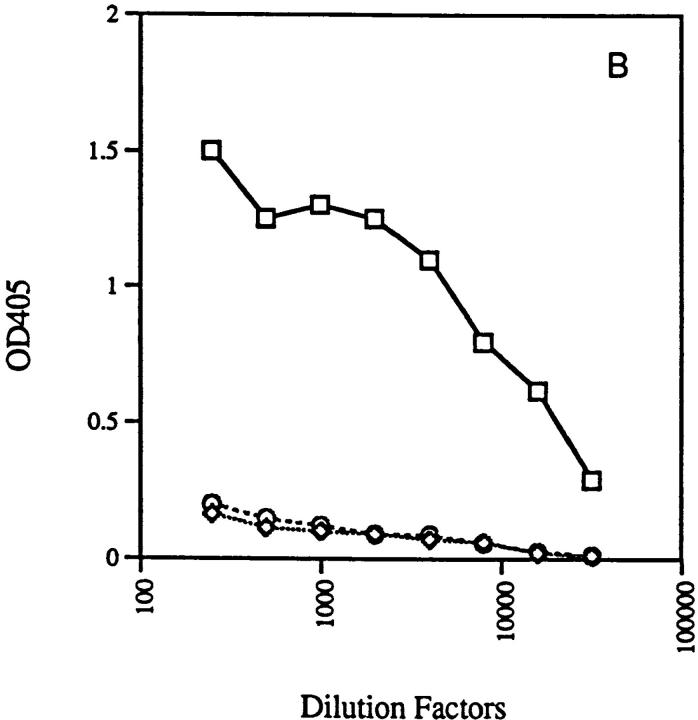
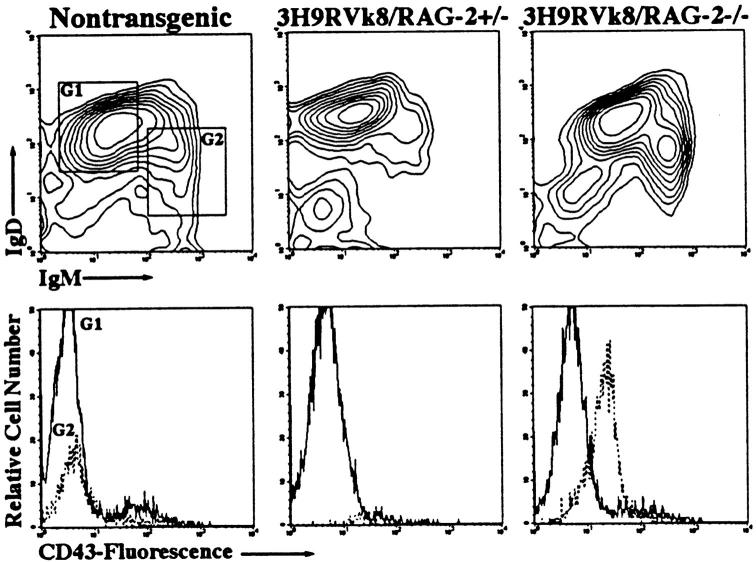
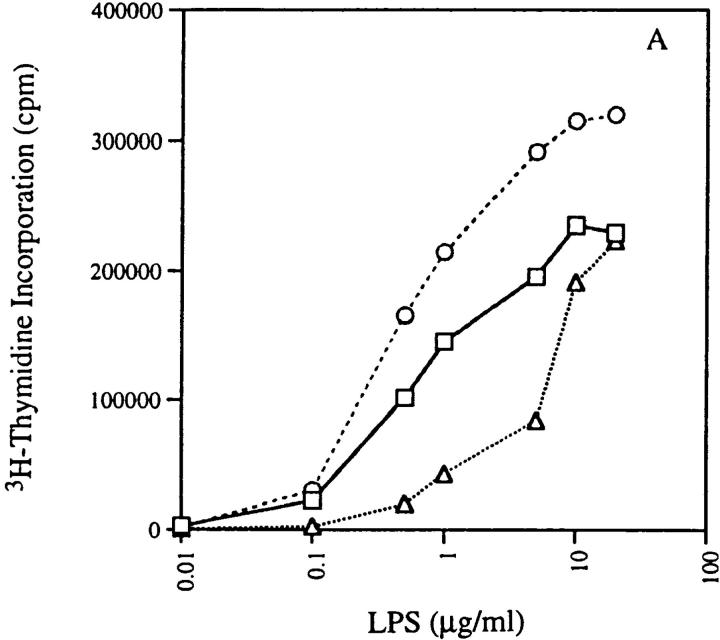
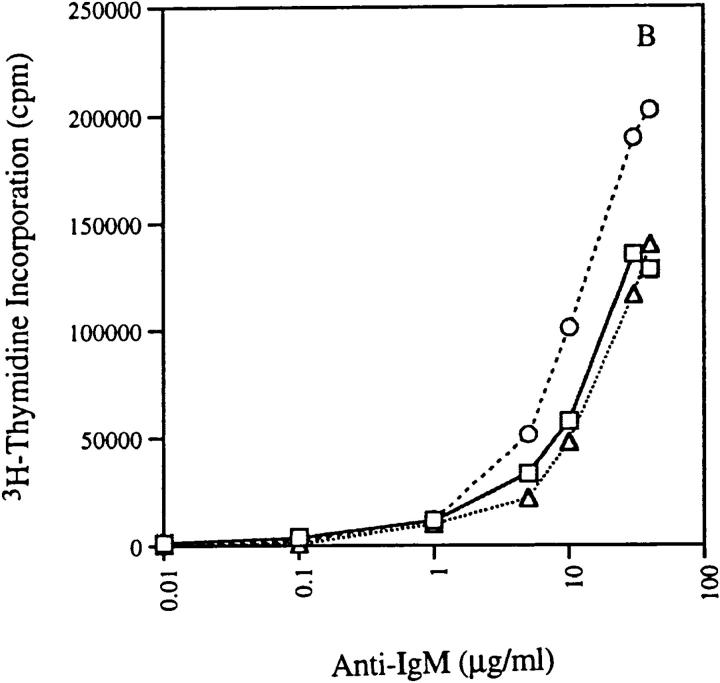
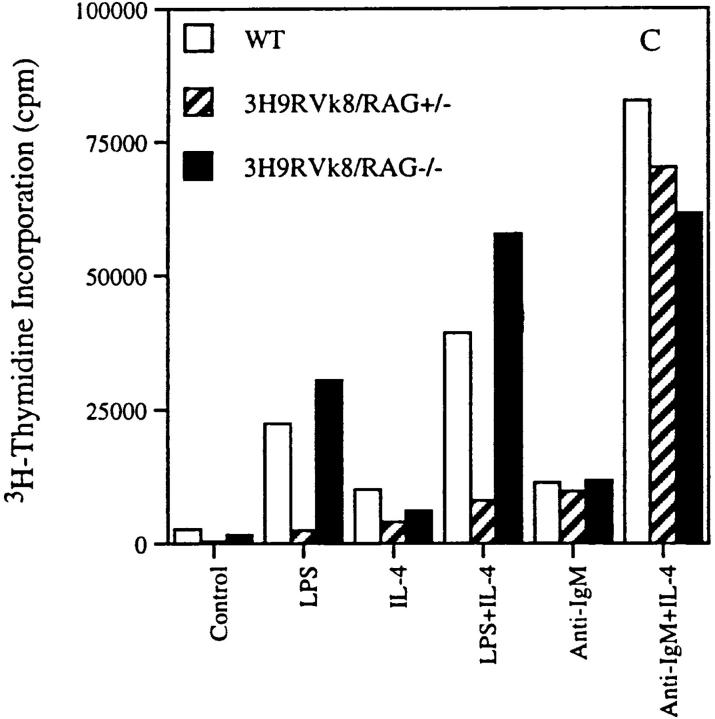
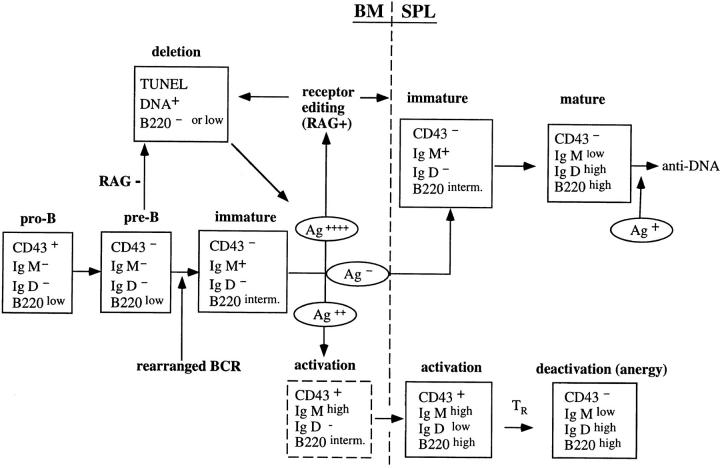
Anti-DNA antibodies are regulated in normal individuals but are found in high concentration in the serum of systemic lupus erythematosus (SLE) patients and the MRL lpr/lpr mouse model of SLE. We previously studied the regulation of anti-double-stranded (ds)DNA and anti-single-stranded (ss)DNA B cells in a nonautoimmune background by generating mice carrying immunoglobulin transgenes coding for anti-DNAs derived from MRL lpr/lpr. Anti-dsDNA B cells undergo receptor editing, but anti-ssDNA B cells seem to be functionally silenced. Here we have investigated how anti-DNA B cells are regulated in recombination- activating gene (RAG)-2-/- mice. In this setting, anti-dsDNA B cells are eliminated by apoptosis in the bone marrow and anti-ssDNA B cells are partially activated.
Figures
References
-
- Tan EM. Antinuclear antibodies: diagnostic markers and clues to the basis of systemic autoimmunity. Pediatr Infect Dis J. 1988;7:S3–S9. - PubMed
-
- Hardin JA. The lupus autoantigens and the pathogenesis of systemic lupus erythematosus. Arthritis Rheum. 1986;29:457–460. - PubMed
-
- Shlomchik MJ, Marshak-Rothstein A, Wolfowicz CB, Rothstein T, Weigert MG. The role of clonal selection and somatic mutation in autoimmunity. Nature. 1987;328:805–811. - PubMed
-
- Craft JE, Hardin JA. Linked sets of antinuclear antibodies: what do they mean? . J Rheumatol. 1987;14:106–109. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
