

Different protein tyrosine kinases are required for B cell antigen receptor-mediated activation of extracellular signal-regulated kinase, c-Jun NH2-terminal kinase 1, and p38 mitogen-activated protein kinase
- PMID: 9763609
- PMCID: PMC2212500
- DOI: 10.1084/jem.188.7.1297
Different protein tyrosine kinases are required for B cell antigen receptor-mediated activation of extracellular signal-regulated kinase, c-Jun NH2-terminal kinase 1, and p38 mitogen-activated protein kinase
Abstract
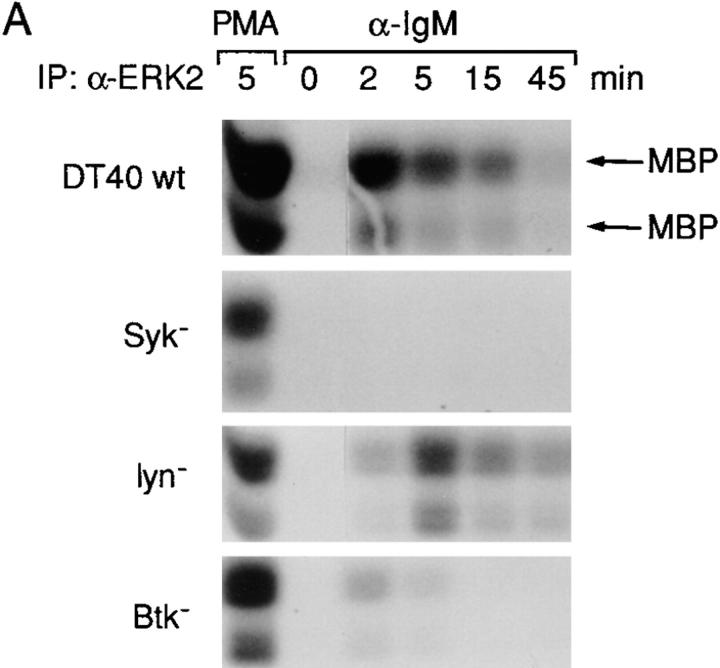
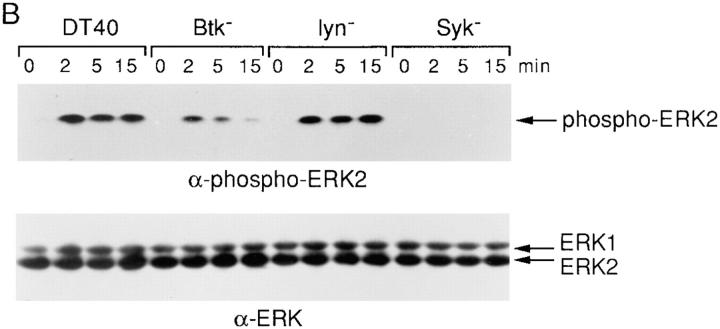
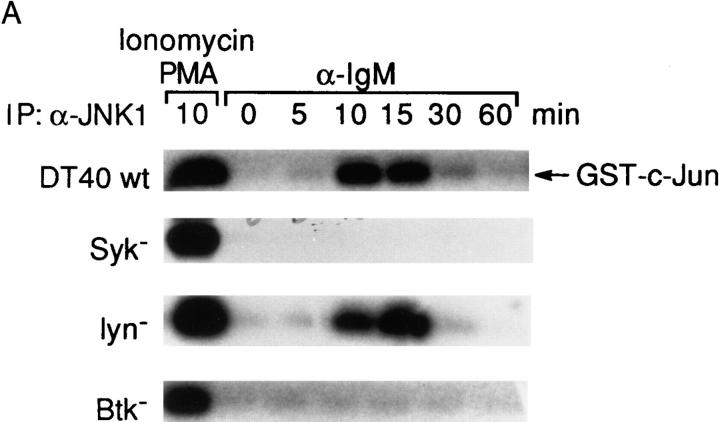
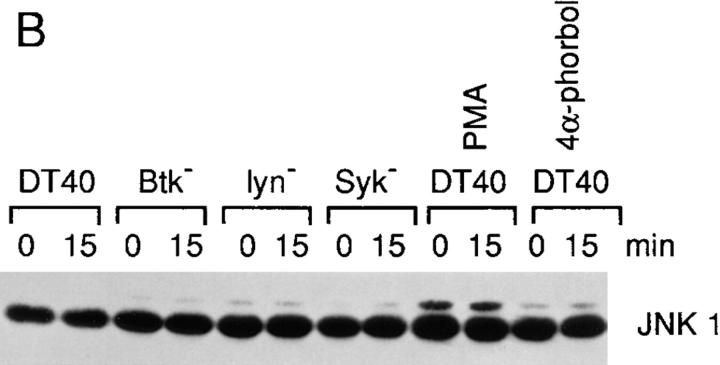
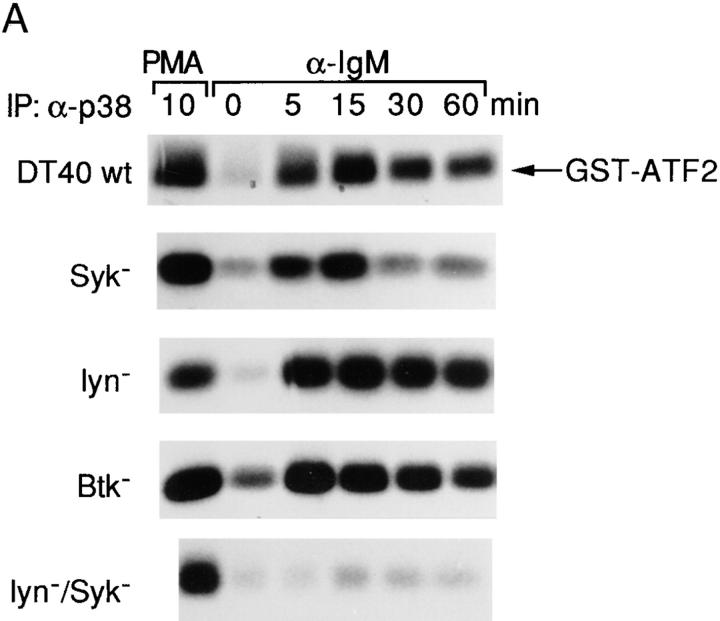
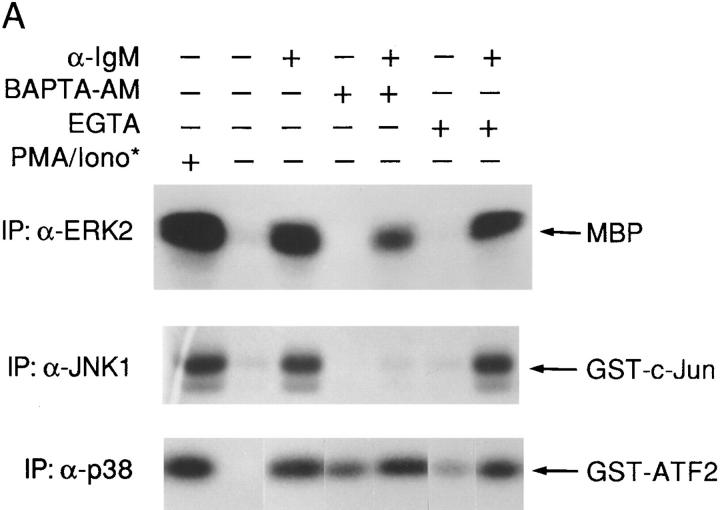
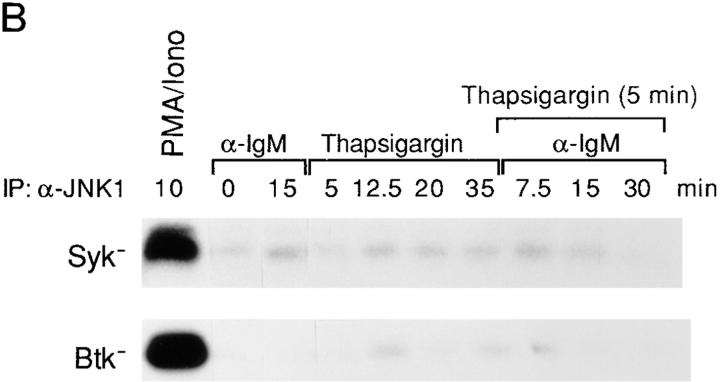
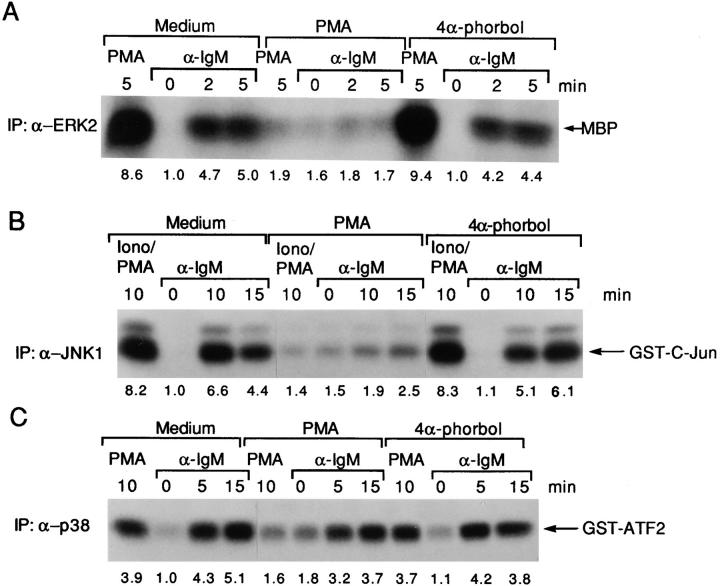
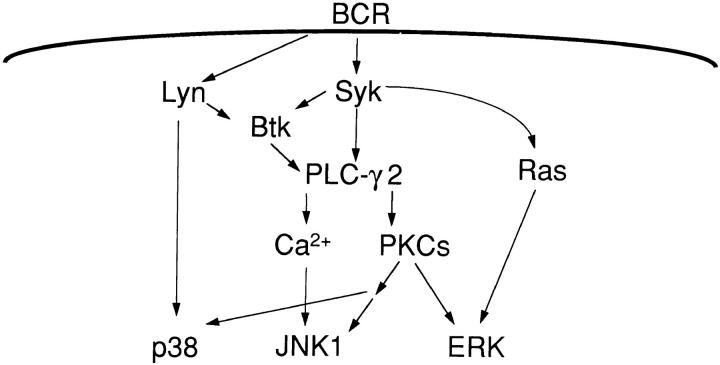
B cell antigen receptor (BCR) cross-linking activates three distinct families of nonreceptor protein tyrosine kinases (PTKs): src-family kinases, Syk, and Btk; these PTKs are responsible for initiating downstream events. BCR cross-linking in the chicken DT40 B cell line also activates three distinct mitogen-activated protein kinases (MAPKs): extracellular signal-regulated kinase (ERK)2, c-jun NH2-terminal kinase (JNK)1, and p38 MAPK. To dissect the functional roles of these PTKs in MAPK signaling, activation of MAPKs was examined in various PTK-deficient DT40 cells. BCR-mediated activation of ERK2, although maintained in Lyn-deficient cells, was abolished in Syk-deficient cells and partially inhibited in Btk-deficient cells, indicating that BCR-mediated ERK2 activation requires Syk and that sustained ERK2 activation requires Btk. BCR-mediated JNK1 activation was maintained in Lyn-deficient cells but abolished in both Syk- and Btk-deficient cells, suggesting that JNK1 is activated via a Syk- and Btk-dependent pathway. Consistent with this, BCR-mediated JNK1 activation was dependent on intracellular calcium and phorbol myristate acetate-sensitive protein kinase Cs. In contrast, BCR-mediated p38 MAPK activation was detected in all three PTK-deficient cells, suggesting that no single PTK is essential. However, BCR-mediated p38 MAPK activation was abolished in Lyn/Syk double deficient cells, demonstrating that either Lyn or Syk alone may be sufficient to activate p38 MAPK. Our data show that BCR-mediated MAPK activation is regulated at the level of the PTKs.
Figures
Similar articles
-
Syk and Bruton's tyrosine kinase are required for B cell antigen receptor-mediated activation of the kinase Akt.J Biol Chem. 1999 Oct 22;274(43):30644-50. doi: 10.1074/jbc.274.43.30644. J Biol Chem. 1999. PMID: 10521450
-
Differential activation of the ERK, JNK, and p38 mitogen-activated protein kinases by CD40 and the B cell antigen receptor.J Immunol. 1996 Oct 15;157(8):3381-90. J Immunol. 1996. PMID: 8871635
-
Syk is required for BCR-mediated activation of p90Rsk, but not p70S6k, via a mitogen-activated protein kinase-independent pathway in B cells.J Biol Chem. 1997 Jul 18;272(29):18200-8. doi: 10.1074/jbc.272.29.18200. J Biol Chem. 1997. PMID: 9218456
-
The Evolving Role of Bruton's Tyrosine Kinase Inhibitors in B Cell Lymphomas.Int J Mol Sci. 2024 Jul 9;25(14):7516. doi: 10.3390/ijms25147516. Int J Mol Sci. 2024. PMID: 39062757 Free PMC article. Review.
-
Syk and pTyr'd: Signaling through the B cell antigen receptor.Biochim Biophys Acta. 2009 Jul;1793(7):1115-27. doi: 10.1016/j.bbamcr.2009.03.004. Epub 2009 Mar 21. Biochim Biophys Acta. 2009. PMID: 19306898 Free PMC article. Review.
Cited by
-
Different sensitivity of germinal center B cell-like diffuse large B cell lymphoma cells towards ibrutinib treatment.Cancer Cell Int. 2014 Apr 3;14(1):32. doi: 10.1186/1475-2867-14-32. Cancer Cell Int. 2014. PMID: 24693884 Free PMC article.
-
Phospholipase Cgamma2 mediates RANKL-stimulated lymph node organogenesis and osteoclastogenesis.J Biol Chem. 2008 Oct 24;283(43):29593-601. doi: 10.1074/jbc.M802493200. Epub 2008 Aug 26. J Biol Chem. 2008. PMID: 18728019 Free PMC article.
-
Tec kinases regulate actin assembly and cytokine expression in LPS-stimulated human neutrophils via JNK activation.Cell Immunol. 2009;258(1):90-7. doi: 10.1016/j.cellimm.2009.03.017. Epub 2009 Apr 23. Cell Immunol. 2009. PMID: 19393603 Free PMC article.
-
TNF receptor-associated factor 3 restrains B-cell receptor signaling in normal and malignant B cells.J Biol Chem. 2021 Jan-Jun;296:100465. doi: 10.1016/j.jbc.2021.100465. Epub 2021 Feb 25. J Biol Chem. 2021. PMID: 33639170 Free PMC article.
-
Inhibitory effect of N-Acyl dopamines on IgE-mediated allergic response in RBL-2H3 cells.Lipids. 2013 Apr;48(4):383-93. doi: 10.1007/s11745-013-3758-6. Epub 2013 Feb 2. Lipids. 2013. PMID: 23377981
References
-
- Reth M. Antigen receptors on B lymphocytes. Annu Rev Immunol. 1992;10:97–121. - PubMed
-
- Bolen JB. Protein tyrosine kinases in the initiation of antigen receptor signaling. Curr Opin Immunol. 1990;7:306–311. - PubMed
-
- Sefton BM, Campbell MA. The role of tyrosine protein phosphorylation in lymphocyte activation. Annu Rev Cell Biol. 1991;7:257–274. - PubMed
-
- Law CL, Craxton A, Otipoby KL, Sidorenko SP, Klaus SJ, Clark EA. Regulation of signalling through B-lymphocyte antigen receptors by cell–cell interaction molecules. Immunol Rev. 1996;153:123–154. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
