

Involvement of PKC-alpha in regulatory volume decrease responses and activation of volume-sensitive chloride channels in human cervical cancer HT-3 cells
- PMID: 9763633
- PMCID: PMC2231219
- DOI: 10.1111/j.1469-7793.1998.435be.x
Involvement of PKC-alpha in regulatory volume decrease responses and activation of volume-sensitive chloride channels in human cervical cancer HT-3 cells
Abstract
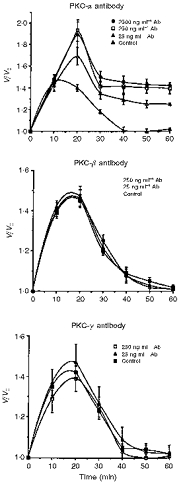
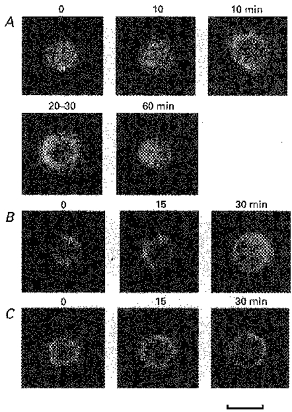
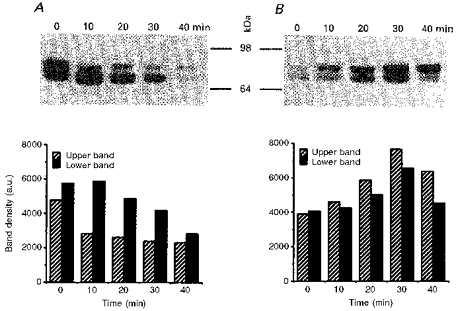
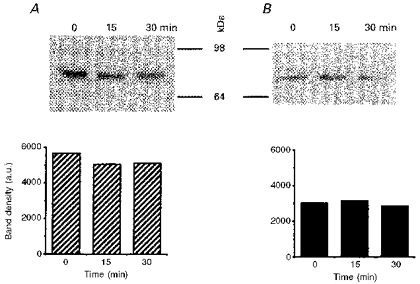
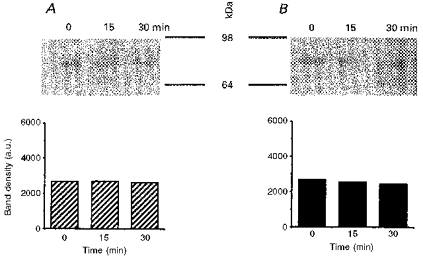
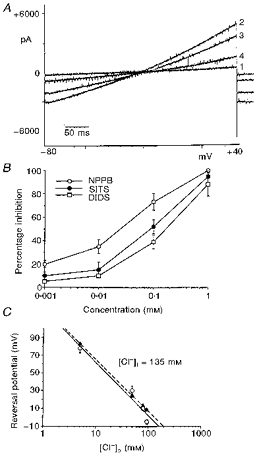
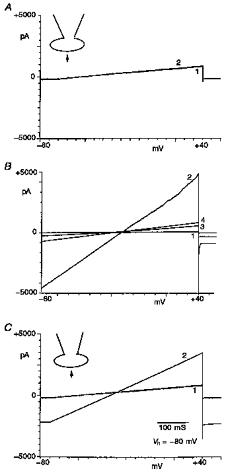
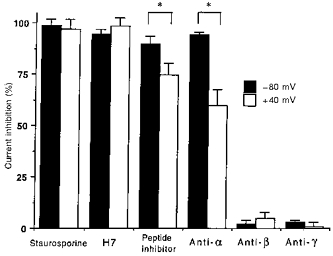
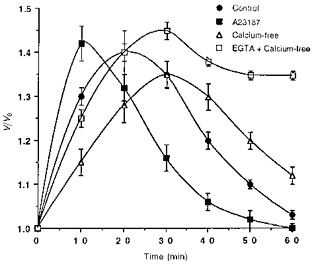
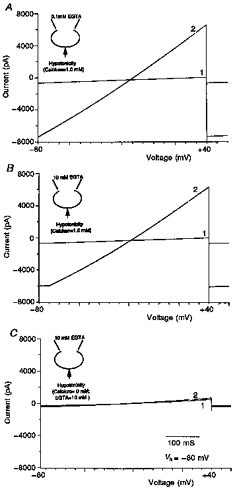
1. The present study was carried out to identify the specific protein kinase C (PKC) isoform involved in regulatory volume decrease (RVD) responses, and to investigate the signal transduction pathways underlying the activation of volume-sensitive chloride channels in human cervical cancer HT-3 cells. The role of Ca2+ in RVD and in the activation of chloride currents was also studied. 2. The time course of RVDs was prolonged by microinjection of PKC-alpha antibody but not by PKC-beta or PKC-gamma antibody, and also by exposure to Ca2+-free medium, in particular when combined with microinjection of EDTA. Immunofluorescence staining showed that hypotonic superfusion evoked the translocation of PKC-alpha to the cell membrane, whereas PKC-beta or PKC-gamma remained unaffected. The translocation of PKC-alpha was observed a few minutes after hypotonic stress, reaching peak intensity at 30 min, and returned to the cytoplasm 60 min after hypotonic exposure. Western blot analyses showed an increased PKC-alpha level in terms of intensity and phosphorylation in the cell membrane, while neither PKC-beta nor PKC-gamma was activated upon hyposmotic challenge. 3. Whole-cell patch-clamp studies demonstrated that neomycin and PKC blockers such as staurosporine and H7 inhibited volume-sensitive chloride currents. The inhibitory effect of neomycin on chloride currents can be reversed by the PKC activator phorbol 12-myristate, 13-acetate (PMA). Moreover, the PKC inhibitor and PKC-alpha antibody, but not PKC-beta or PKC-gamma antibody, significantly attenuated the chloride currents. The activation of volume-sensitive chloride currents were insensitive to the changes of intracellular Ca2+ but required the presence of extracellular Ca2+. 4. Our results suggest the involvement of PKC-alpha and extracellular Ca2+ in RVD responses and the activation of volume-sensitive chloride channels in HT-3 cells.
Figures
References
-
- Borner C, Filipuzzi I, Wartmann M, Eppenberger U, Fabbro D. Biosynthesis and post-translational modifications of protein kinase C in human breast cancer cells. Journal of Biological Chemistry. 1989;264:13902–13909. - PubMed
-
- Chang WC, Ning CC, Lin MT, Huang JD. Epidermal growth factor enhances a microsomal 12-lipoxygenase activity in A431 cells. Journal of Biological Chemistry. 1992;267:3657–3666. - PubMed
-
- Chou CY, Shen MR, Chen TM, Huang KE. Volume-activated taurine transport is differentially activated in human cervical cancer HT-3 cells, but not activated in HPV-immortalized Z 183A and normal cervical epithelial cells. Clinical and Experimental Pharmacology and Physiology. 1997;24:935–939. - PubMed
-
- Chou CY, Shen MR, Wu SN. Volume-sensitive chloride channels associated with human cervical carcinogenesis. Cancer Research. 1995;55:6077–6083. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous