

Coronavirus transcription early in infection
- PMID: 9765389
- PMCID: PMC110261
- DOI: 10.1128/JVI.72.11.8517-8524.1998
Coronavirus transcription early in infection
Abstract
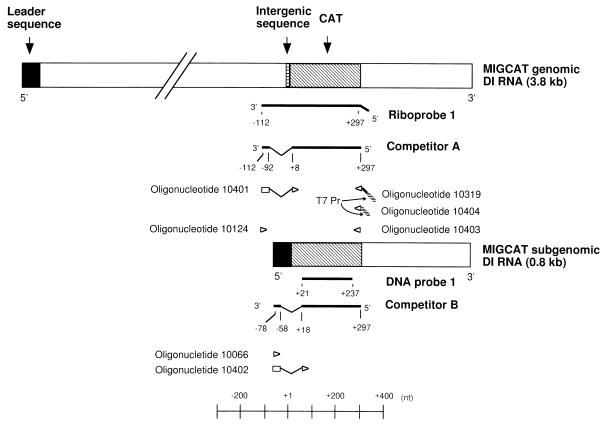
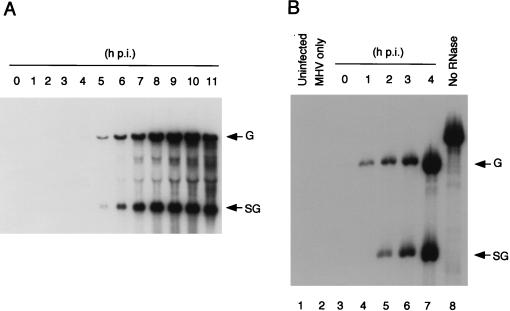
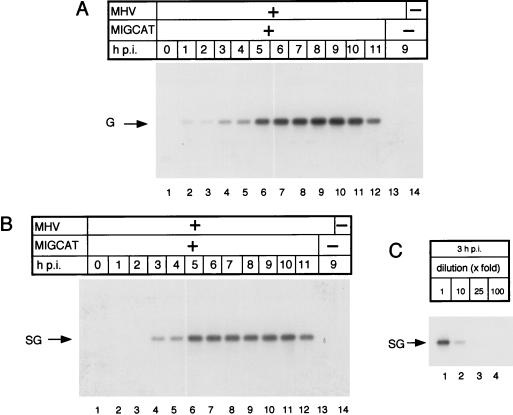
We studied the accumulation kinetics of murine coronavirus mouse hepatitis virus (MHV) RNAs early in infection by using cloned MHV defective interfering (DI) RNA that contained an intergenic sequence from which subgenomic DI RNA is synthesized in MHV-infected cells. Genomic DI RNA and subgenomic DI RNA accumulated at a constant ratio from 3 to 11 h postinfection (p.i.) in the cells infected with MHV-containing DI particles. Earlier, at 1 h p.i., this ratio was not constant; only genomic DI RNA accumulated, indicating that MHV RNA replication, but not MHV RNA transcription, was active during the first hour of MHV infection. Negative-strand genomic DI RNA and negative-strand subgenomic DI RNA were first detectable at 1 and 3 h p.i., respectively, and the amounts of both RNAs increased gradually until 6 h p.i. These data showed that at 2 h p.i., subgenomic DI RNA was undergoing synthesis in the cells in which negative-strand subgenomic DI RNA was undetectable. These data, therefore, signify that negative-strand genomic DI RNA, but not negative-strand subgenomic DI RNA, was an active template for subgenomic DI RNA synthesis early in infection.
Figures

References
-
- Hirano N, Fujiwara K, Hino S, Matsumoto M. Replication and plaque formation of mouse hepatitis virus (MHV-2) in mouse cell line DBT culture. Arch Gesamte Virusforsch. 1974;44:298–302. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
