

Circular intermediates of recombinant adeno-associated virus have defined structural characteristics responsible for long-term episomal persistence in muscle tissue
- PMID: 9765395
- PMCID: PMC110267
- DOI: 10.1128/JVI.72.11.8568-8577.1998
Circular intermediates of recombinant adeno-associated virus have defined structural characteristics responsible for long-term episomal persistence in muscle tissue
Erratum in
- J Virol 1999 Jan;73(1):861
Abstract
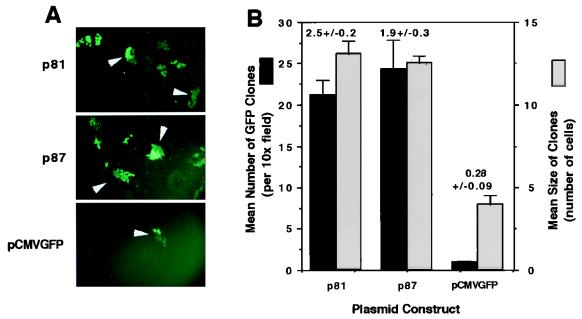
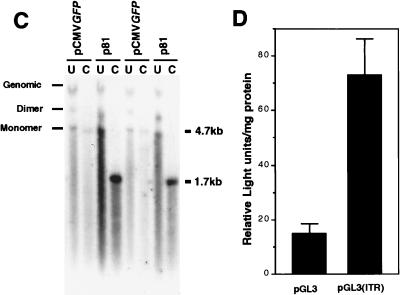
Adeno-associated viral (AAV) vectors have demonstrated great utility for long-term gene expression in muscle tissue. However, the mechanisms by which recombinant AAV (rAAV) genomes persist in muscle tissue remain unclear. Using a recombinant shuttle vector, we have demonstrated that circularized rAAV intermediates impart episomal persistence to rAAV genomes in muscle tissue. The majority of circular intermediates had a consistent head-to-tail configuration consisting of monomer genomes which slowly converted to large multimers of >12 kbp by 80 days postinfection. Importantly, long-term transgene expression was associated with prolonged (80-day) episomal persistence of these circular intermediates. Structural features of these circular intermediates responsible for increased persistence included a DNA element encompassing two viral inverted terminal repeats (ITRs) in a head-to-tail orientation, which confers a 10-fold increase in the stability of DNA following incorporation into plasmid-based vectors and transfection into HeLa cells. These studies suggest that certain structural characteristics of AAV circular intermediates may explain long-term episomal persistence with this vector. Such information may also aid in the development of nonviral gene delivery systems with increased efficiency.
Figures
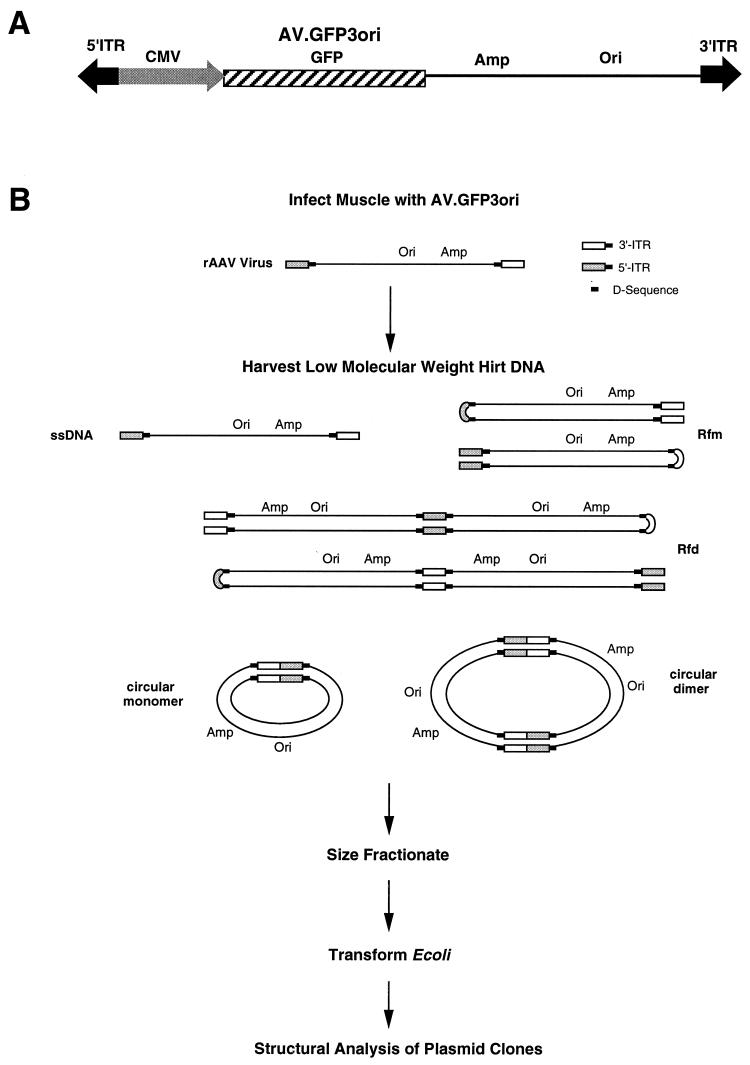

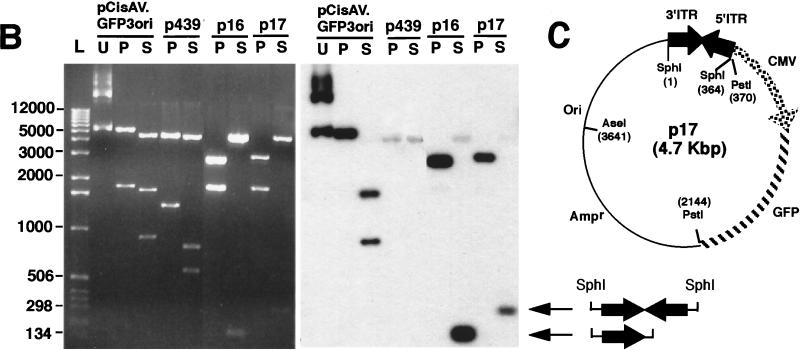
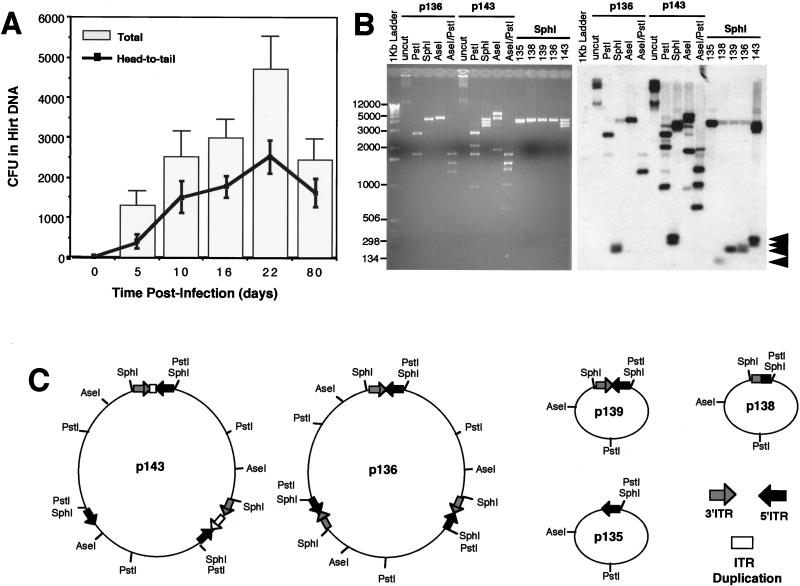
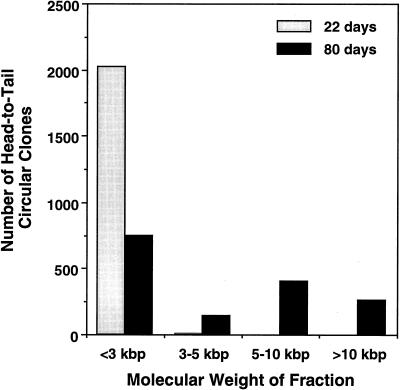
References
-
- Bennett J, Duan D, Engelhardt J F, Maguire A M. Real-time, noninvasive in vivo assessment of adeno-associated virus-mediated retinal transduction. Investig Ophthalmol Visual Sci. 1997;38:2857–2863. - PubMed
-
- Clark K R, Sferra T J, Johnson P R. Recombinant adeno-associated viral vectors mediate long-term transgene expression in muscle. Hum Gene Ther. 1997;8:659–669. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
